Die Reoviridae bilden die größte Familie von unbehüllten Viren mit doppelsträngiger RNA (dsRNA). Zusammen mit den Birnaviridae und Partitiviridae sind sie die einzigen Viren, deren dsRNA-Genom segmentiert vorliegt. Zur Familie gehören so wichtige Erreger wie das Blauzungenvirus, die humanen Rotaviren oder das Colorado-Zeckenfieber-Virus. Der Name der Familie ist ein Akronym aus „respiratory, enteric, orphan“, da man der Ansicht war, dass Mitglieder dieser taxonomischen Gruppe nur Darmerkrankungen (enteric), Atemwegserkrankungen (respiratory) oder keine Erkrankung (orphan) hervorrufen würden. Tatsächlich wurden die Reoviridae bei vielen Säugetieren gefunden, ebenso bei Reptilien, Fischen, Krustentieren und Insekten. Drei Gattungen der Familie (Fijivirus, Phytoreovirus und Oryzavirus) umfassen Erreger bei Pflanzen (Pflanzenviren) und Pilzen (Mykoviren).
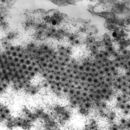
Die reifen Virusteilchenen (Virionen) der Reoviridae sind unbehüllte, ikosaedrische Kapside mit einem Durchmesser von etwa 60–80 nm. Charakteristisch für die meisten Gattungen der Familie (mit Ausnahme der Gattung Cypovirus) ist der Aufbau des Virions aus zwei ineinander geschachtelte und eng wechselwirkende Kapside (T=13), die wiederum eine innere, regelmäßige Corestruktur umgeben. Die so aus drei Lagen von Proteinen aufgebauten Virionen zeigen daher im Elektronenmikroskop eine sehr typische, dickwandige Struktur mit regelmäßigen, speichenförmigen Verdickungen. Von dieser Speichenstruktur abgeleitet erhielt die Gattung Rotavirus (lat. rota: Rad) ihren Namen. Diese Speichenstruktur entsteht durch 132 Öffnungen des äußeren Kapsids, die in ebenso viele Kanäle des inneren Kapsids münden. Diese Kanäle, die bis an das innere Core reichen, lassen eine Diffusion von Ionen in das Virion zu.
Dadurch, dass das innere Core und das innere Kapsid für die korrekte Verpackung der RNA-Segmente verantwortlich sind, kann das äußere Kapsid eine relativ große Variabilität der Oberflächenstrukturen aufweisen ohne die Stabilität des Virions und die Vollständigkeit des Genoms zu beeinträchtigen. Daher zeigen sich bei vielen Reoviren zahlreiche Spezies und Serotypen.
In der inneren Coreschale befindet sich ein Molekül der viralen RNA-Polymerase und Guanyltransferase. Bei einigen Gattungen sind Myristyl-Reste (Tetradecyl-Reste) kovalent an das Kapsid gebunden. Bei den Gattungen Rotavirus, Orbivirus und Coltivirus kommen unreife Virionen vor, die Reste einer Lipidhülle besitzen. Bei diesen Gattungen geschieht das Verlassen der Zelle durch Knospung an der Zellmembran (Orbivirus, Coltivirus) oder der Membran des Endoplasmatischen Retikulums (ER) (Rotavirus). Durch einen enzymatischen Reifungsschritt verlieren die Virionen die Lipidhülle wieder. Bei einigen Gattungen wird das äußere Kapsid durch wirtseigene Proteasen (z. B. Trypsin, Chymotrypsin) verändert, wodurch sogenannte „infectious subviral particles“ (ISVPs) entstehen. Nur die ISVPs sind infektiös und der Übergang von unreifen Virionen zu ISVPs spielt bei diesen Viren eine entscheidende Rolle in der Krankheitsentstehung und der unterschiedlichen Erscheinungsform der Virionen im Darm, Speichel oder Blutserum.
Das Genom besteht aus einer doppelsträngigen RNA, die je nach Gattung in 10 bis 12 Segmente unterteilt ist. Die Molmasse der Segmente reicht von 0,2 bis 3 × 106 Da. Die virale mRNA der Reoviridae besitzt keinen Poly-A-Schwanz. Bei einigen Virusspezies findet man kurze, einzelsträngige RNA-Oligonukleotide in der inneren Coreschale.
Mit Stand November 2018 gliedern sich nach Maßgabe des ICTV die Reoviriridae wie folgt (neben den Typusspezies ist nur eine Auswahl weiterer Spezies angegeben):[3]
Das folgende Kladogramm gibt vereinfacht die Verwandtschaftsbeziehungen wieder, wie sie im 9. Report des International Committee on Taxonomy of Viruses (ICTV, 2011) und von MacLachlan/Dubovi (2016);[4][11] sowie fast übereinstimmend von Belhouchet et al. (2010)[12] vorgeschlagen wurde:
Reoviridae SedoreovirinaeMimoreovirus (MPrV)
Orbivirus (SCRV, CHUV, AHSV, BTV, KEMV, TRBV, EHDV)
Phytoreovirus (RDV)
Seadornavirus (LNV, KDV, BAV)
Cardoreovirus (ESRV)
Rotavirus (RVA, RVB, RVC)
Orthoreovirus (MRV)
Oryzavirus (RRSV)
Dinovernavirus (APRV)
Coltivirus (EYAV, CTFV)
Für einige der Gattungen sind die abgekürzten Speziesnamen angegeben, diese umfassen jeweils einen oder mehrere der untersuchten zugehörigen Stämme (englisch strains).
In älteren Arbeiten gibt es noch gewisse Abweichungen (z. B. bei Attoui et al. (2006)[13] bei Quito-Avila (2011)[14] und bei Ke et al. (2010).[15]
Das ICTV hat mit der Master Species List #35 vom März 2020 die Reoviridae dem neu geschaffenen Phylum Duplornaviricota zugeordnet.[16] Eine Kladogramm findet sich bei Picornavirales §ICTV Master Species List #35.
Die Reoviridae bilden die größte Familie von unbehüllten Viren mit doppelsträngiger RNA (dsRNA). Zusammen mit den Birnaviridae und Partitiviridae sind sie die einzigen Viren, deren dsRNA-Genom segmentiert vorliegt. Zur Familie gehören so wichtige Erreger wie das Blauzungenvirus, die humanen Rotaviren oder das Colorado-Zeckenfieber-Virus. Der Name der Familie ist ein Akronym aus „respiratory, enteric, orphan“, da man der Ansicht war, dass Mitglieder dieser taxonomischen Gruppe nur Darmerkrankungen (enteric), Atemwegserkrankungen (respiratory) oder keine Erkrankung (orphan) hervorrufen würden. Tatsächlich wurden die Reoviridae bei vielen Säugetieren gefunden, ebenso bei Reptilien, Fischen, Krustentieren und Insekten. Drei Gattungen der Familie (Fijivirus, Phytoreovirus und Oryzavirus) umfassen Erreger bei Pflanzen (Pflanzenviren) und Pilzen (Mykoviren).




