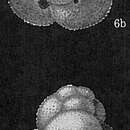
Globigerinoides ruber (d'Orbigny 1839) was originally described under the name Globigerina rubra by d'Orbigny (1839) and later designated as the type species of by (Cushman 1927). The test or shell of G. ruber is multi-chambered and trochospirally coiled; that is, all chambers are visible on the spiral side, while only those of the last-formed whorl are visible on the opposite, or umbilical, side (Bé et al., 1969). The early chambers of the test have a distinct reddish coloration which is the origin of the species name rubra or ruber. The chambers are subspherical in shape and increase in size as new chambers are added to pre-existing chambers during the life of the cell (Brummer et al. 1987). There are three chambers in the last whorl and a total of 15 chambers in the typical adult test (Bé et al. 1969). The terminal chamber may be a normal shape (normalform), or it may be somewhat smaller than the preceding chambers (diminutive), or it may be bubble-like and extend to the umbilical region (bulla-like) (Brummer et al., 1987). The sutures between the chambers are depressed and radial in orientation.
The shape of the primary aperture, or opening into each chamber, is a high, semicircular arch that is bordered by a narrow rim (Banner and Blow 1960). This aperture is interiomarginal in location (that is, it occurs at the suture between the distal wall and preceding coil), and umbilical in position (it opens from the final chamber directly into the umbilicus on the umbilical side of the test) (Banner and Blow 1960). Paired, symmetrical supplementary apertures that are sutural in position develop later in ontogeny and are located above sutures of earlier chambers on the spiral side of the test (Brummer et al. 1987, Schiebel and Hemleben 2017). These supplementary apertures are shaped like semi-circular arches and are also bordered by narrow rims (Banner and Blow 1960).
The shell is made of secreted calcite and is bilamellar in construction (Schliebel & Hemleben, 2017). The test wall ranges in thickness from <5µm in the juvenile portion of the test to 20 µm in the final chambers of the last whorl (Bé et al. 1969). The surface of the test is coarsely spinose with a honeycomb (cancellate) texture (Hemleben et al. 1991). The pores in the test wall average 4.4 µm in diameter, occur at a density of five pores per 25 µm2 of test surface area, resulting in a test porosity of 11.6% (Bé et al. 1969). Unlike some other spinose planktonic foraminiferans, G. ruber produces little or no calcite crust, a calcite veneer that coats the test, at the end of their life cycle (Caron et al., 1990).
A dense array of thin, round spines project from the chamber surfaces of Globigerinoides ruber (Type 2 of Hemleben et al. 1991). The spines are formed from needlelike crystals of calcite that are anchored in spine collars or bases located on the surface of the shell between the pores (Saito et al. 1976). The spines are approximately 10 times longer than the test is wide, with the average test width of G. ruber measuring 560 µm in diameter (Schiebel and Hemleben 2017). Spines are shed during gametogenesis, the process of gamete formation (Caron et al. 1990). The release of gametes in G. ruber appears to be synchronized with lunar cycles and occurs with a half-synodic lunar periodicity (Schiebel and Hemleben 2005).
Live individuals surround the shell with a spherical cytoplasmic capsule that is comprised of a network of granuloreticulose pseudopodia, anastomosing arrays of fine cytoplasmic strands, strung between the dense array of calcite spines (Bé 1982, Bé et al. 1977, Hemleben et al. 1989, Schiebel and Hemleben 2017). This three-dimensional halo of pseudopodia and spines that surrounds the shell can be further subdivided into three zones: (1) an outer zone formed by the tips of the spines and fine strands of granulose reticulopodia; (2) an intermediate zone of denser granuloreticulose pseudopodia; and (3) an inner zone populated by commensal microalgae (Pyrocystis noctiluca and P. robusta) and denser granuloreticulose pseudopodia (Hemleben and Spindler 1983, Schiebel and Hemleben 2017).
When alive, individuals of G. ruber host intracellular dinoflagellate symbionts that have been identified as belonging to the species Pelagodinium beii (Fujiki et al. 2014, Siano et al. 2010). These endosymbionts may supplement the diet of the host foraminiferan with excess photosynthetic products; however, recent studies of photosymbiosis in Globigerinoides (=Trilobatus) sacculifer indicate that the symbionts contribute very little to the daily nutrition of the cell and may serve mostly as an emergency food source, as well as a supplemental energy source during reproduction (Takagi et al. 2018).
Globigerinoides ruber is mixotrophic as it can derive nutrition from both the photosynthetic products of its autotrophic algal symbionts, and by feeding on suspended food particles which are engulfed by the cell through phagocytosis (Takagi et al. 2018). Although this species is omnivorous and feeds primarily on copepods in laboratory experiments (Caron et al. 1990), a diversity of food sources have been observed in its digestive vacuoles, including diatoms, dinoflagellates and pieces of animal tissue (Bé 1982).
Living populations of G. ruber inhabit the sunlit waters of the photic zone in the open ocean from 0- to 30-m water depths (Hemleben & Spindler, 1983). Globigerinoides ruber appears to spend its entire life cycle within the uppermost layers of the water column and does not sink to deeper depths during prior to gametogenesis, as has been observed in other species of spinose planktic foraminiferans (Hemleben & Spindler, 1983).
In laboratory experiments, G. ruber has been shown to tolerate water temperatures ranging from 14-32 ºC, with optimum growth rates achieved at 23 ºC (Bijma et al. 1990). Likewise, this species can tolerate salinities that range from 22-49 ppt, with optimum growth rates occurring at 35.5 ppt (Bijma et al. 1990).
Parker (1962) differentiated three distinct phenotypic variants of G. ruber that she considered to result from growth under different environmental regimes. Other authors, working on both modern and fossil specimens, have subdivided G. ruber into two morphotypes, a “pink” morphotype and a “white” morphotype, and Wang (2000) further subdivided the “white” morphotype in two recognizably distinct forms based on their stable isotopic compositions of oxygen and carbon. Recent molecular analyses have shown that a broad interpretation of the Globigerina ruber morphospecies can be subdivided into at least four species-level subclades, represented by the genotypes Ia+Ib (white), “pink”, IIa+IIb, and Globigerinoides conglobatus (“white”) (Aurahs et al. 2011, Darling and Wade 2008, Kuroyanagi et al. 2008, Ujiié and Lipps 2009). It has been recommended that species epithet ruber be restricted to specimens that exhibit the “pink” morphotype or chromotype (Aurahs et al. 2011).
Globigerinoides ruber has a cosmopolitan distribution in the world ocean at subtropical-tropical latitudes (Hemleben et al. 1989, Schiebel and Hemleben 2017). The geologic range of the Globigerinoides ruber lineage has been estimated to extend back 7.5 million years to the Late Miocene (Aze et al. 2011), an estimate that closely corresponds to a 6.3 million year old molecular clock estimate of the divergence time between the G. ruber (“pink” chromotype) and its sister clade, the clade comprised of the unnamed Type I + Type II genotypes (Aurahs et al. 2011).
The types of Globigerinoides ruber(d’Orbigny, 1839) are deposited in the Paleontology Collection of the Muséum National d’Histoire Naturelle (http://www.mnhn.fr) in Paris, France. A lectotype was designated from d’Orbigny’s original syntype suite by Banner and Blow (1960: p. 20). A photomicrograph of the lectotype, along with scanning electron micrographs of topotypic specimens, were published by Le Calvez (1977: Pl. 5, figs. 1-4 (topotypes), 5 (lectotype)). The museum number for the lectotype is MNHN.F.FO302 {MHNH, 2018a). Seven paralectotypes are housed under the museum number MNHN.F.FO246 (MHNH, 2018b).
The original collecting locality is cited as shore sands from an unspecified location on the island of Cuba (d’Orbigny, 1839; Le Calvez, 1977).