Comprehensive Description
provided by Smithsonian Contributions to Zoology
Abyssochrysos melvilli (Schepman)
Argyropeza melvilli Schepman, 1909:170, pl. 12, fig. 1 [holotype: Zoologisches Museum, Amsterdam, 2915; type-locality: Sulu Sea, 535 m, Siboga Expedition, Sta 102].
Abyssochrysos melvilli (Schepman).—Barnard, 1963b: 141–144.x
Abyssochrysos tomlini Barnard, 1963b: 143–144, fig. 27f [lectotype, herein designated: larger of two syntypes, South African Museum, A9101; type-locality: 24 mi off Cape Natal (Durban) at 801 m, South Africa].
Argyropezai suvaensis Ladd, 1977:15–16, pl. 1: fig. 3; pl. 21: fig. 8 [holotype: USNM 174966; paratype: USNM 175062; type-locality: Suva, Fiji, early Miocene].
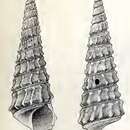
DESCRIPTION.—Shell (Figure 7): Shell thin, elongate, turreted, and consisting of 13–16 whorls in adults. Adult mean shell length 36 mm, mean width 9 mm wide, length-width ratio of 4:1, and with apical angle of 15–19 degrees. Whorls weakly inflated, pedunculate at their bases, sculptured with 17–18 strong, sinuous, biangulate axial ribs and numerous, minute, sinuous, axial growth lines that cross over the ribs. Each axial rib with a blunt nodule at top and slightly larger one at bottom. Top row of modules connected by weak spiral cord. Base of each whorl with fine but distinct spiral cord adajacent to suture. Suture strongly impressed. Axial sculpture not strong and sometimes lacking on body whorl. Base of body whorl concave, with two spiral cords at periphery and 5–7 weaker spiral cords on concave portion. Protoconch smooth, one and one-half whorls, about one-seventh the length of the shell. Columella concave, smooth, and with thin callous. Outer lip very sinuous, thin. Weak, anterior siphonal shelf present, no umbilicus. Periostracum thin, glossy, and of brownish green to brownish yellow color.
Animal: Animal about 6–7 whorls, having prominent, thick snout that is bilobed at tip with slit-like mouth between lobes. Cephalic tentacles thick, about one-half width of snout. No eyes. Foot moderately short, with glandular lobes on propodium. Circular attachment scar for operculum at posterior dorsal surface of foot. Mantle skirt smooth, apparently lacking papillae at edge, and folded at right dorsal center, forming respiratory cavity. Mantle cavity deep, about two and one-half whorls and two-thirds the length of the animal. Thin osphradium and long, wide bipectinate ctenidium begin close behind mantle skirt and extend length of left portion of mantle cavity. No hypobranchial glands seen. Pallial gonoducts closed, thin, adjacent to right portion of columellar muscle. Large, dorsal, penis-like process arises from mantle under right edge of mantle skirt. Smaller papilla to the right of “penis” also arises from mantle skirt. “Penis” and associated papilla with numerous circular folds. Base of “penis” flattened where connected to mantle and with tubular connection to gonoduct. Intestine long and thin, filled with small ovoid fecal pellets that are composed of fine sediment. Anal opening behind and dorsal to “penis,” about 2.5 mm from mantle edge. Buccal mass relatively large and muscular. Nerve ring and associated ganglia tightly concentrated. Two chitinous, semi-lunarshaped jaws present in sides of anterior buccal cavity. Jaws tan, small in relation to buccal mass, about 0.35 mm long, and with weakly serrated edges; composed of numerous plates that are characteristically subrhomboidal but occasionally pentagonal. Operculum (Figure 7e) chitinous, thin, ovate, and paucispiral with a subcentral sunken nucleus. Edges of previous opercular whorls slightly laminate. Growth lines oblique, microscopic. Attachment scar of foot muscle almost round, occupying central portion of operculum. Color golden brown, but darker around circumference.
Radula (Figure 8): Radula robust, long (radular ribbon 3.25 mm in length, 0.50 mm wide and with 108 rows of teeth in animal 12 mm long). Radula identical to that of A. melanoides except rachidian tooth thinner at center.
MATERIAL EXAMINED.—2 specimens, lectotype and paralectotype (South African Museum A-9101), dredged in 801 m by R. S. Pieter Faure off Cape Natal, South Africa; 1 specimen (USNM 237535), dredged by 956 m off Cabra Is., Lubang, Philippines; 1 specimen (USNM 244970), 490 m off Malavatuan Is., W. Luzon, Philippines; 1 specimen (USNM 239417), 1074 m, off North Is., Buton Strait, Celebes, Indonesia; 1 specimen (USNM 239426), 983 m off Tg Lamulu, Gulf of Boni, Celebes, Indonesia; 1 specimen (USNM 258507), 866 m off Sipadan Is., Sibuko Bay, Borneo, Indonesia; 1 specimen (USNM 239341), 1472 m off Doworra Is., Moluccas, Indonesia; 1 specimen (USNM 239392), 945 m off North Is., Buton Strait, Celebes, Indonesia; 1 specimen (USNM 239343), 1472 m off Doworra Is., Moluccas, Indonesia.
Discussion
Before considering the inclusion of the Abyssochrysidae in the Loxonematacea, a word should be said about the state of the material upon which some of the evidence is based. The animals of Abyssochrysos melanoides and A. melvilli had been dried in their shells for many years and were poorly preserved. Many organs, such as the stomach, were too compressed to examine even after reconstitution. None of the material was suitable for histological study. Although I was able to study the mantle cavity and its associated organs, I could not ascertain unequivocally the presence of a duct or lumen through the pallial gonoducts. This structure had been compressed and the internal ducts were in a collapsed state. Nevertheless, I believe my observations relative to the closed state of the pallial gonoducts and their connection to the mantle process or “penis” are correct, as explained below.
ANATOMY.—There are a number of objections that may be raised about the transfer of the family Abyssochrysidae to the superfamily Loxonematacea, a group thought to be extinct since the Upper Jurassic (Knight et al, 1960:1315). The most obvious one is the fact that we have no way of knowing what the soft parts of the animals in this extinct group were like, and must rely solely on shell characters for speculation about their phylogenetic relationships. Although the shells of the two Abyssochrysos species are very similar to those of the Paleozoic loxonemataceans (see Figure 11), there is the possibility that this is due to convergence and does not indicate relationship. For example, the shells and periostracum of Abyssochrysos species are similar to those of species in the genera Melanoides A. Adams, Stenomelania Fischer and Namratua Abbott of the large fresh-water family Thiaridae Troschel (= Melaniidae Latreille). Indeed, Tomlin (1927) named Abyssochrysos melanoides in reference to this resemblance. There is a striking likeness of sculpture between Melanoides admirabilis E. A. Smith, 1880, and A. melanoides, although the aperture of the former is more attenuate. One can find other examples of sculptural resemblance among the thiarids, but similarity between the Abyssochrysidae and Thiaridae is clearly attributable to sculptural convergence. The apertures of the latter group are more elongate and fusiform; moreover, the thiarids are anatomically cerithiaceans.
The presence of closed pallial gonoducts and large penis-like organs in the species of Abyssochrysos excludes them from the superfamily Cerithiacea because the distinguishing anatomical characters of members of the superfamily are open pallial gonoducts and the aphallic condition of males; there are a few minor exceptions to this among the Pleuroceridae, which will be discussed below. One might argue that the Caecidae Gray, which traditionally has been placed in the Cerithiacea Fleming (Thiele, 1931: Wenz, 1939) is an exception to this, but modern workers no longer consider the Caecidae to be among the cerithiaceans. Fretter and Graham (1962:625) noted that the anatomical features of Caecum were highly unusual for a cerithiacean and suggested that the caecids were accidentally assigned to the superfamily merely on the basis of shell structure. Moore (1962:699) has shown that the anatomy of the Caecidae places them in the superfamily Rissoaceae Gray.
Other cerithiacean families with species that resemble the general shell physiognomy of the Abyssochrysidae are the Cerithiidae Fleming, Syrnolopsidae Bourguignat, Turritelidae Woodward, and Potamididae H. and A. Adams. All members of these large families are characterized by their open pallial gonoducts. The Cerithiidae and Potamididae have shells with distinct anterior canals. Johansson (1953:17) considered the open pallial gonoducts of Cerithiacea to be a primitive trait. He also noted (1956:386) that the closed pallial gonoducts of the Rissoidae appear to be derived from open slits. Fretter (1951:583) and Fretter and Graham (1962: 359) did not agree that the open ducts in the Cerithiacea were primitive characters and suggested that the open condition is correlated with a long, narrow mantle cavity and is thus a secondary and advantageous trait. If the Abyssochrysidae are indeed primitive, the presence of closed pallial gonoducts in this group would tend to support the latter point of view.
Another large group of mesogastropods to which the Abyssochrysidae may be related in the Rissoacea Gray. As I remarked earlier, Golikov and Starabogatov (1975:219) provisionally assigned the family to the Rissoacea on the basis of shell characters. There is sculptural similarity to some freshwater rissoaceans, such as Oncomelania Gredler, 1881, and also to some marine rissoaceans like Truncatella Risso, 1826, and Rissoina Orbigny, 1847. This similarity is superficial; most marine rissoids are tiny animals and their shells lack an anterior siphonal canal. No described rissoacean radula resembles those of the Abyssochrysos species. Although little is known of the anatomy of many groups assigned to the superfamily Rissoacea, those that have been studied have females with closed pallial oviducts and males with cephalic penis and closed gonoducts. Exceptions are to be found in the subfamily Rissoininae Stoliczka, in which the condition of the male pallial gonoduct may be open or closed (Ponder, 1967:209), and in the Bythiniidae, in which the penis of Bithynia tentaculata is of pallial origin (Fretter and Graham, 1962:350). In some rissoids the presence of a penis in females, as well as in males, has been noted by Thiriot-Quievreux (1977:781), who considered the condition due to successive hermaphroditism. Thus, although there are a few anatomical similarities between members of the Rissoacea and Abyssochrysidae, there is not enough evidence to consider the latter group as rissoaceans.
The only other marine families that approach the general shell shape of the Abyssochrysidae are the Terebridae Mörch and Pyramidellidae Gray. The former are advanced toxoglossans and have a narrow aperture and a distinct twisted columella with a lower plait, while the latter are opisthobranchs, easily distinguished by their heterostrophic nuclear whorls and detorted anatomy.
Abyssochrysos shells are sculptured like those of extinct loxonemataceans in the family Pseudozygopleuridae. An interesting feature in Abyssochrysos is that the axial scultpure does not necessarily follow the growth lines of the shell and is thus not colabral sculpture. This is not the pattern in cerithid shells but is true for some loxonemataceans (Robert Linsley, pers. comm.). Fossil loxonemataceans have consistently thicker shells and straighter columellas than the abyssochrysids. The labial sinus noted by Knight et al. (1960) in the shells of some fossil loxonematids has traditionally been interpreted as an anal sinus and, if in a central position, as an indication of a zygobranchous organization. Although a labial sinus does not occur in Abyssochrysos species, the presence of a pallial penis could account for the sinus in extinct forms of loxonemataceans. Thus, a labial sinus in fossils does not necessarily indicate the presence of an exhalant siphon and associated anus.
Perhaps the most unusual feature of the abyssochrysid snails is the presence of the large pallial process and its associated papilla. Although I interpret this structure as a penis, it may also be considered to have a number of other functions. It may be a right pallial tentacle such as exists in some rissoaceans like Cingula Fleming, 1828 (Fretter and Graham, 1962:112) or in Valvata Müller 1774 of the family Valvatidae Thompson (Fretter and Graham, 1962:621). In these groups the pallial tentacle functions as a ciliated extension of the exhalant siphon. In Abyssochrysos the pallial process is very wide, thick, and not at all tentaculate in appearance. Moreover, there is an apparent axial lumen present and the proximal part of the process is attached to the inner surface of the mantle, a short distance behind the mantle edge. There is also an internal connection to the pallial gonoduct; accordingly, it is doubtful that the process is homologous to the papillae or tentacles of the mantle skirt.
Barnard (1963b:141) regarded the pallial process of Abyssochrysos as an extension of the oviduct. Only two of the animals he examined had this structure and he considered them to be females. The other four lacked the process and were regarded as males. I find Barnard's interpretation unlikely because I know of no other prosobranchs in which the oviduct overhangs the mantle skirt. It is difficult to explain the functional significance of such a condition. I think it is simpler to consider this structure a penis and to regard the four animals lacking it as females. All of the animals of A. melanoides I examined had the process, although in a few it was much reduced. The one animal of A. melvilli that I studied also had the structure. Because I am not certain that the mantle process is a penis, I cannot unequivocally identify the sex of the animals I studied. There are two possibilities: (1) all the animals I saw were males; (2) males as well as females have the pallial process. The fact that the process is reduced in some animals indicates that it may be a senescent feature or that the Abyssochrysidae could be successive hermaphrodites. Barnard (1963b:141) thought that the papilla next to the penis was an anal papilla, but the anus is located far behind the papilla and has no apparent association with it. I believe it best to regard this structure as an accessory copulatory papilla.
Another possible interpretation of the pallial process is that it is an ovipositor. This is also doubtful because all other known ovipositors in mesogastropods are pedal structures and, from a functional view point, the position of an ovipositor above the head seems unlikely.
In my view, the interpretation of this structure as a penis is most acceptable. The penis is of pedal origin in most mesogastropods, but a penis of pallial origin, although unusual, is not unprecedented. Several examples occur in the superfamily Viviparacea among members of the Pilidae Connolly and Viviparidae Gray. In Viviparus the right cephalic tentacle is modified to function as a penis (Fretter and Graham, 1962:350). It is in the freshwater Pilidae that one finds structures analogous, though probably not homologous, to the condition observed in the Abyssochrysidae. Andrews (1964:123–124) has shown that in the Pilidae the penis arises from right portion of the mantle and that it occurs in both males and females. The penis in this group differs from that of the Abyssochrysidae by having an elaborate penis sheath associated with it. Andrews (1966:134) suggested that the penis in the Pilidae may have originated from a pallial tentacle such as that of Valvata.
Other mesogastropods having a penis of mantle origin are in the family Pleuroceridae Fischer, superfamily Cerithiacea. Moore (1898:191, pl. 14:figs. 45–46) described and figured a penis-like organ arising from the mantle and connected by a duct to the middle of the male pallial gonoduct in Typhobia horei, a member of the subfamily Typhobiinae Moore, endemic to lake Tanganyika. Moore (1898:191) considered this organ to be an eversible penis but did not think it was homologous with the penis of the Pilidae (cited as Ampullariae). Morrison (1954:373) later commented that the Tiphobiinae were the only “Melanian” snails that possess any intromittent organ structure whatsoever. In Typhobia, the gonoduct is open only at its lower extremity. Thus the Typhobiinae differ markedly from most other cerithiaceans, a group characterized by aphallic males and open pallial gonoducts.
Members of the genera Cleopatra Troschel and Paludomus Swainson, in the subfamily Paludominae also differ from “standard” pleurocerid reproductive anatomy. In Cleopatra the female pallial gonoduct is closed, according to Starmuhlner (1969:181), but his drawing of this organ (fig. 259) is equivocal enough to render his interpretation of a “closed” gonoduct questionable. In Paludomus, as in Typhobia, the female pallial gonoduct is open only at its lower extremity (Seshaiya, 1934:207). The male pallial gonoduct is unusual in being swollen and closed except at the terminal portion where it becomes an open ciliated slit. A nonmuscular flagellar penis runs forward in the connective tissue of the mantle and lies on the rectal side of the terminal portion of the gonoduct (Seshaiya, 1934:203–204). This situation is similar to that observed in Abyssochrysos. Morrison (1954:369), however, discounted Seshaiya's interpretation, and noted that the penis was non-muscular and could not function as an intromittent organ. It is thus clear that the reproductive anatomy of the Paludominae, although still not clearly understood, is exceptional and does not resemble that of other cerithiaceans.
Fretter and Graham (1962:350) noted that in the Bithyniidae Troschel, superfamily Rissoacea, Bithynia tentaculata has a penis of pallial origin, although it appears superficially to emerge from the head.
Radula: The radulae of the two Abyssochrysos species (Figures 5,8) are nearly identical and although definitely taenioglossate, are much different from all other taenioglossate radulae. The distinctive rachidian and lateral teeth are not similar to those of any other mesogastropods. A survey of numerous prosobranch radulae in Troschel (1856–1863), Sars (1878), Schepman (1909), and in a number of papers by Ponder (1965, 1966, 1967) and Ponder and Yoo (1976) illustrating radulae of unusual rissoaceans revealed none similar to Abyssochrysos.
SYSTEMATIC PLACEMENT.—The characters derived from the shell physiognomy, anatomy, and radulae of the two Abyssochrysos species do not fit the taxonomic limits of any Recent prosobranch superfamily. The bathyal habitat of these snails is also unusual. Similarly, there are no Cenozoic prosobranch groups to which the Abyssochrysidae may be assigned. One is thus confronted with the prospect of proposing a new superfamily. On the other hand, members of several families in the Paleozoic superfamily Loxonematacea Koken, 1889 have shell physiognomies that are strikingly like those of the Abyssochrysidae (Figure 11). The Paleozoic loxonemataceans were abundant in the Permian (Knight, 1930) and occur as fossils as late as the Upper Jurassic. There are three families in this group that closely resemble the Abyssochrysidae: the Paleozygopleuridae Horny, 1955, the Pseudozygopleuridae Knight, 1930, and the Zygopleuridae Wenz, 1938. They are characterized by species with turreted shells that are frequently ribbed and have smooth protoconchs of one and one-half whorls. Inner shell layers are not nacreous. Early members of the Loxonematacea have a slight sinus that tends to move toward the suture and become shallower in genera from later periods, but this feature is not present in all genera. According to Knight et al. (1960), the Pseudozygopleuridae arose in the Mississippean Period and comprised many genera and species. The Zygopleuridae were most common in the Triassic and Upper Jurassic. The loxonemataceans were common components of Paleozoic shallow-water faunas of inland seas. Numerous fossils of this group occur in shales (Knight, 1930:23) and some pseudozygopleurids are thought to have lived on muddy bottoms (David Schindel, pers. comm.). Although the loxonemataceans are thought to have disappeared after the Jurassic, it is interesting to note that early Miocene fossils of Abyssochrysos melvilli have been found in Fiji (Ladd, 1977).
ECOLOGY.—There are only two known living species of Abyssochrysos, both of which are uncommon and confined to bathyal zones of continental shelfs. Although the deep sea is no longer regarded as a refugium for an ancient fauna, (Clarke, 1962: Knudsen, 1970) both the abyssal and bathyal zones of the deep sea have provided habitats for other unusual mollusks that had their origins in the Paleozoic such as pleurotomaracean snails, monoplacphorans, and many archaic bivalves and scaphopods. Other invertebrate groups as well as some vertebrates of ancient lineage survive only in the deep sea. Parker (1962:286) suggested that many of the shelf-bottom faunas migrated down the slope during the Paleozoic and early Mesozoic times in response to competition. Menzies, George, and Rowe (1973:359) stated that most deep-sea endemic genera and families are derived from warm-water ancestors of the Mesozoic and Tertiary. They also pointed out that deep-sea animals have most likely been derived from shallow-water communities living on fine sediments. Most archaic types are found today on shelf or archibenthal depths rather than in the abyssal depths. Fossil loxonematids have thicker, heavier shells than Abyssochrysos species and lived in shallow seas. The thinner shells of the Abyssochrysidae are indicative of their bathyal habitat and are, in this respect, typical of deep-sea snails. Thus, it is not unreasonable to suggest that a few members of the Loxonematacea have persisted in this habitat. To date there are no records of Abyssochrysos from the abyssal zone.
Conclusions
The superfamily Loxonematacea is considered to be ancestral to the Cerithiacea (Fretter and Graham, 1962); both have been assigned to the order Caenogastropoda (=Mesogastropoda + Neogastropoda Wenz) by Knight et al (1960). The gross anatomy and taenioglossate radulae of the Abyssochrysidae are indicative of simple mesogastropods related to the superfamilies Viviparacea, Valvatacea, Rissoacea, and Cerithiacea. Rather than create a new superfamily to accommodate them and violate the principle of parsimony, I am provisionally recognizing the Abyssochrysidae as a relict family of loxonematid snails probably most closely related to the families Zygopleuridae and Pseudozygopleuridae. The collection of more specimens with well-preserved soft parts is necessary for any further speculation about their exact relationships within the Prosobranchia.
- bibliographic citation
- Houbrick, Richard S. 1979. "Classification and systematic relationships of the Abyssochrysidae, a relict family of bathyal snails (Prosobranchia: Gastropoda)." Smithsonian Contributions to Zoology. 1-21. https://doi.org/10.5479/si.00810282.290