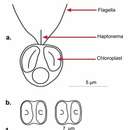
Els haptòfits (Haptophyta) és un grup d'algues unicel·lulars. Els seus cloroplasts es pigmenten de forma semblant a la dels heteroconts (com en les crisofícies), però es diferencien en la resta de l'estructura cel·lular, de manera que pot ser que siguin una línia evolutiva separada en la que els cloroplasts es deriven d'endosimbiosis similars.
Les cèl·lules tenen típicament dos flagels lleugerament desiguals, que són llisos, i un únic orgànul denominat haptonema, que se superficialment similar a un flagel però que es diferencia del mateix en el conjunt de microtúbuls i en el seu ús. Els mitocondris tenen crestes tubulars.
Els hiptòfits més coneguts són els cocolitòfors (ordres dels cocolitals i isocrisidals), que tenen un exoesquelet de plaques calcàries denominades cocolitos. Constitueixen el fitoplàncton marí més abundant, especialment en mar obert i és extremadament abundant com a microfòssil. Altres haptòfits planctònics inclouen Chrysochromulina i Prymnesium, que periòdicament produeixen floracions algals marines tòxiques, i Phaeocystis que produeix floracions que poden generar una escuma desagradable que s'acumula sovint a les platges. Estudis moleculars i morfològics divideixen els haptòfits en cinc ordres.
Els haptòfits (Haptophyta) és un grup d'algues unicel·lulars. Els seus cloroplasts es pigmenten de forma semblant a la dels heteroconts (com en les crisofícies), però es diferencien en la resta de l'estructura cel·lular, de manera que pot ser que siguin una línia evolutiva separada en la que els cloroplasts es deriven d'endosimbiosis similars.
Les cèl·lules tenen típicament dos flagels lleugerament desiguals, que són llisos, i un únic orgànul denominat haptonema, que se superficialment similar a un flagel però que es diferencia del mateix en el conjunt de microtúbuls i en el seu ús. Els mitocondris tenen crestes tubulars.
Els hiptòfits més coneguts són els cocolitòfors (ordres dels cocolitals i isocrisidals), que tenen un exoesquelet de plaques calcàries denominades cocolitos. Constitueixen el fitoplàncton marí més abundant, especialment en mar obert i és extremadament abundant com a microfòssil. Altres haptòfits planctònics inclouen Chrysochromulina i Prymnesium, que periòdicament produeixen floracions algals marines tòxiques, i Phaeocystis que produeix floracions que poden generar una escuma desagradable que s'acumula sovint a les platges. Estudis moleculars i morfològics divideixen els haptòfits en cinc ordres.
Haptophyta (případně Haptophytina, dříve též Prymnesiophyta) je skupina převážně mořských jednobuněčných bičíkovců s fotosyntetickou schopností. Jako u naprosté většiny ostatních fotoautotrofů říše Chromista vznikl jejich plastid sekundární endosymbiózou z ruduchy.
Postavení skupiny na fylogenetickém stromě eukaryot není dosud plně vyjasněno. Některé současné fylogenetické studie ho kladou buď do podříše Hacrobia[1], jiné přirozenost hacrobií neuznávají a haptophyta spolu s centrohelidními slunivkami považují za sesterský klad ke kladu SAR[2] nebo je ponechávají jako eukaryota incertae sedis[3][4].
Mají dva bičíky a navíc haptonemu (připomíná třetí bičík). Povrch bičíků je pokryt mastigonematy, uvnitř nich je typická tzv. axonematická struktura. Naopak haptonema žádnou axonemu uvnitř nemá. Na povrchu buněk jsou nad periplastem často kladeny tenké ploché šupinky, buď z polysacharidů, nebo navíc z vápenatých sloučenin. Druhá jmenovaná chemická struktura je důvodem vzniku tzv. kokolitů, tedy zbytků těchto řas, které se ukládají na dně moří ve velkém množství.[5]
K původnímu rozsahu této skupiny, tvořeném tzv. prymnesiofyty (odtud dřívější synonymum haptofyt – Prymnesiophyta), přibyly díky molekulárním analýzám dvě menší skupiny, nyní samostatné třídy.[1] Poslední z nich, tzv. rappemonády, byly objeveny sekvenováním mořských vzorků teprve v r. 2010.[6]
Někdy se ke stejnému kmeni přiřazuje i třída centrohelidních slunivek, takto rozšířený kmen se nazývá Haptista a haptofyta jsou ponížena podkmen Haptophytina.
Aktuální (2015) klasifikace:[1]
Haptophyta (případně Haptophytina, dříve též Prymnesiophyta) je skupina převážně mořských jednobuněčných bičíkovců s fotosyntetickou schopností. Jako u naprosté většiny ostatních fotoautotrofů říše Chromista vznikl jejich plastid sekundární endosymbiózou z ruduchy.
Postavení skupiny na fylogenetickém stromě eukaryot není dosud plně vyjasněno. Některé současné fylogenetické studie ho kladou buď do podříše Hacrobia, jiné přirozenost hacrobií neuznávají a haptophyta spolu s centrohelidními slunivkami považují za sesterský klad ke kladu SAR nebo je ponechávají jako eukaryota incertae sedis.
Die Haptophyta (früher auch Prymnesiophyta genannt) sind überwiegend marin-planktonisch lebende, komplexe einzellige Algen. Die Gruppe gehört innerhalb der Diaphoretickes entweder zu den Harosa (alias Sar) oder zu den Hacrobia.
Die gelben, gelbbraunen oder braunen Chloroplasten enthalten Chlorophyll a und c, β-Carotin und Xanthophylle. Als Reservestoffe werden Chrysolaminarin, Öl und selten auch Paramylon (auch als Paramylum bezeichnet) abgelagert. Die Chloroplasten befinden sich in einer Falte des Endoplasmatischen Reticulums. Die Chloroplasten ähneln denen der Heterokonten, der übrige Zellaufbau weicht jedoch von diesen ab, weshalb die Haptophyta heute systematisch von den Heterokonta getrennt werden.
Begeißelte Zellen verfügen über zwei meist gleich lange oder nur wenig unterschiedlich lange Geißeln (isokonte Begeißelung). Die Geißeln sind nicht wie bei den Heterokonta mit Flimmerhaaren besetzt, sondern mit submikroskopischen Schüppchen oder Knoten. Im Gegensatz zu den Heterokonten besitzen die Geißeln auch keine geschwollene Basis (Geißelschwellung).
Neben den Geißeln besitzt jede Zelle ein Haptonema. Es ist ein fadenförmiges Anhängsel, das der Anheftung dient. Der Aufbau ähnelt oberflächlich dem einer Geißel, im Querschnitt befinden sich jedoch 6 oder 7 sichelförmig angeordnete Mikrotubuli.
Die Zellen sind außen mit Schuppen aus Polysacchariden (meist Cellulose), die in Golgi-Vesikeln gebildet und anschließend an der Zelloberfläche abgelagert werden, besetzt. Bei der Ordnung Coccolithales sind die äußeren Schuppen verkalkt und werden Coccolithen genannt. Die Funktion der Coccolithen ist bislang nur unzureichend bekannt. Vermutet werden u. a. Fraßschutz, Auftriebsregulation, Lichtzuführung oder Kalzifikation zur Unterstützung der Photosynthese. Teilweise ist auch ein Augenfleck, wie bei den Goldalgen und den Euglenozoa vorhanden.
Obwohl Vertreter der monadalen Organisationsstufe überwiegen, kommen auch capsale, kokkale und trichale Vertreter vor. Manche Arten besitzen einen heteromorphen Generationswechsel, d. h. die verschiedenen Generationen sind äußerlich gut unterscheidbar. Bei diesen Kalkalgen wechselt eine diploide (mit doppelten Chromosomensatz) Generation, bei der die Algen Flagellen besitzen und im Plankton leben, mit einem haploiden Stadium (mit einfachem Chromosomensatz), in dem sie benthisch, d. h. auf dem Meeresboden, leben.
Nur wenige Arten leben im Süßwasser, die meisten Arten leben planktonisch im Meer. Sie können in sehr großen Mengen auftreten und einen wesentlichen Teil des Meeresplanktons bilden. Somit spielen die planktonischen Haptophyten eine bedeutende Rolle als Primärproduzenten im Meer. Einige Arten sind kosmopolitisch verbreitet, die Mehrheit kommt allerdings in den Meeren der gemäßigten Breiten vor. In besonders großen Mengen tritt die kosmopolitisch verbreitete Coccolithales-Art Emiliania huxleyi auf. Sie zählt zu den wichtigsten Produzenten von biogenem Calciumcarbonat und kann großräumige Algenblüten verursachen.
Haptophyten treten auch als Symbionten auf, so als Zooxanthellen in Acantharien[1].
Ihre systematische Stellung war lange unklar, sie wurde mittlerweile durch molekularbiologische Methoden weitgehend überarbeitet und geklärt[2][3]. Die Abteilung wird in folgende Klassen eingeteilt:
Die Haptophyta (früher auch Prymnesiophyta genannt) sind überwiegend marin-planktonisch lebende, komplexe einzellige Algen. Die Gruppe gehört innerhalb der Diaphoretickes entweder zu den Harosa (alias Sar) oder zu den Hacrobia.
De Haptophyta sünd en Algenstamm. De Chloroplasten sünd ähnelk farvlich as de Heterokonten.
De Haptophyta sünd Protisten. Dat is en egene Riek un kene Deeren oder Planten. De Zellen hebbt meest en beten unglike Flagella.
Haptofite ili haptofiti (latinski: Haptophyta ili Prymnesiophyta) su skupina, koljeno jednoćelijskih algi s oko 500 vrsta, uglavnom morskih, za koje je karakteristično prisustvo dva biča, haptoneme (kontraktilni dodatak koji površno nalikuje bičevima) i golih ćelija ili prekrivenih karbonatnim pločama.[1][2][3] Uključuje poznate kokolitoforide koje imaju egzoskelet od krečnjačkih plakove ili kokolita, zvanih kokosfera i čine najbogatiju morsku površinu fitoplanktona, posebno u otvorenom moru, a izuzetno su obilne i kao mikrofosili. Neki haptofiti periodično proizvode cvjetanje algi otrovne morske vode ili stvaraju neugodnu pjenu koja se često nakuplja na plažama.
Haptofite su važna sastavnica morskog fitoplanktona i zbog visoke koncentracije imaju važnu ulogu kao primarni proizvođači u tim ekosistemima. U tropskim vodama ih je obilnije, a čine oko 45% fitoplanktona u srednjim širinama južnog Atlantika. Njihova brojnost smanjuje se prema umjerenim širinama, iako su neki haptofiti još uvijek i u polarnim vodama. Nekoliko vrsta živi u slatkoj vodi ili u tlu. Mnoge su vrste su simbioti drugih organizama, naprimjer, tvore asocijacije na radiolarijama.
Neke vrste povremeno proizvode cvetanje algi i otrovne morske vode ili stvaraju neugodnu pjenu koja se često nakuplja na plažama. Neke vrste izbacuju snažne egzotoksine koji uzrokuju visoku smrtnost riba i mehkušaca.
Haptofite su jednoćelijske, mada se ponekad grupiraju u kolonije ili niti. Životni ciklus obično sadrži jednu fazu, ali se u nekim slučajevima izmjenjuju faze sa i bez biča. Bičasta faza je jednoćelijska i planktonska, dok je nebičasta obično kolonijalna i bentoska. Bentoski oblici mogu biti nitasti, kokoidni ili palmeloidni [4] Haptofite su uglavnom fotosintetski organizmi, mada su neki članovi miksotrofi ili heterotrofi. Mnoge vrste su fagofrofne i u citoplazmi imaju vezikule koje probavljaju progutani materijal. Nisu selektivni u pogledu prehrambenih materija koje unose u vezikule i mogu se sastojati i od organskog otpada i bakterija i drugih malih organizama.[5]
Haptofitne ćelije obično imaju dve jednake ili blago neravne i glatke flagele (biča). Izuzetak je red Pavlovales, gdje je jedan od bičeva duži od drugog, a prekriveni su malim dlačicama ili ljuskicama. Haptofiti mogu plivati naizmjenično tako da se flagele usmjere prema naprijed ili prema stražnjoj strani tijela. Kretanje je brzo i često mijenjaju smjer. Često imaju organelu zvan haptonema, koja je površinski slična biču, ali se od njega razlikuje u skupu mikrotubula, kao i u upotrebi. Ime joj dolazi od grčkih korijena hapsis (=dodir) i nema (=nit). Ovisno o vrsti, haptonema može biti dugačka ili kratka, te biti spiralno namotana ili ne. Nalazi se između dvaju flagela i ne kreće se poput ovih, ali može se smanjiti i istezati, a njena bi funkcija bila fiksacija na supstrat, interveniranje u pokretu ili podržavanje sposobnosti fagotrofije kod nekih haptofita. Haptonema se sastoji od sedam jednostavnih mikrotubula okruženih membranom endoplazmatskog retikuluma.
Iako su uključeni neki oblici sa golim ćelijama (Pavlovales), većina ima ćelije prekrivene plakovima ili ljuskicama, tipski eliptičnim. Organske pločice potiču iz Golgijevog aparata i često su umotane u sluz. Ploče mogu biti mineralizirane, a sastoje se uglavnom od ugljikohidrata, ali mogu se i kalcificirati, jer su ploče sa silicijem vrlo rijetke.
Vrste koje sa kalcificiranim plakovima poznate su kao kokolitoforide s. Imaju egzoskelet karbonatnih ploča zvani "kokolit" i predstavljaju najčešću morski fitoplankton, posebno u otvorenom moru. Arhitektura kokolitofora vrlo je raznolika, često sa složenim ukrasima bodlji i prirubnica, a poznate su vrste eliptičnog oblika, peterokutne, zvijezdaste, diskoidne itd. Kokolitofore su uključene u redove Coccolithales i Isochrysidales i izuzetno su brojne kao mikrofosili. Poznate su od karbona, od prije otprilike 300 miliona godina, av bilo ijh je jako puno tokom jure.
Haptofite su u svakoj ćeliji obično imaju jedan do četiri hloroplasta izduženog diskoidnog oblika sa pirenoidom u centru hloroplasta ili kao ispupčenje na jednoj strani. TilakoidI su grupisani tri po tri i nemaju perifernu tilakoidnu lamelu. Hloroplasti su okruženi sa četiri membrane, pri čemu je najudaljenija pokrivena s ribosomima i povezana s jedarnom membranom, tvoreći hloroplastni endoplazmatski reticulum. Kao fotosintetski pigmenti imaju hlorofile a, c1, c2 i c3 (potonji je u Prymnesiophyceae). Kao dodatni pigmentI, pored β-karotena imaju ksantofile tipa dianinoksantina, diatoksantina i fukoksantina. Kao rezervne supstance imaju hrizolaminarin, paramil i kapi lipida, koji se formiraju u vakuolama izvan hloroplasta. Smatra se da su hloroplasti haptofita izvedeni iz sekundarne endosimbioze sa Rhodophyta. Hloroplasti su pigmentirani na način sličan onima kod Ochrophyta (heterokontofitas, kao u Chrysophyceae), ali se razlikuju u ostatku ćelijske strukture, tako da mogu biti zasebna evolucijskaa linija u kojoj su hloroplasti izvedeni iz neovisne endosimbioze (u prvom redu Prymnesiophyceae).[6] Iz ćelijske površine potiču pseudopodije i filipodi koji mogu biti linearni ili razgranati. Mnogi su fagotrofi i posljedično u citoplazmi imaju hranjive vacuole u kojima probavljaju bakterije i druge male organizme. Oni takođe imaju veliki Golgijev aparat koji se nalazi na prednjem kraju ćelije, a u nekim slučajevima i kontraktilne vakuole. Nekoliko rodova imaju očnu mrlju osjetljivu na svjetlost, naprimjer, Pavlova i Diacronema.[7] Očna mrlja smještena je unutar ili u blizini hloroplasta i sastoji se od kapljica lipida.[8] Mitohondrije imaju cjevaste grebene. Haptofiti se obično razmnožavaju aseksualno, prostom diobom i otvorenom mitozom . U nekim grupama može biti često seksualno razmnožavanje.[9] Neke vrste su pronađene s diplohaplontnim ciklusom, u kojem se flagelirani diploidni stadij izmjenjuje s filamentoznim haploidnim . Diploidna faza je planktonska, dok je haploidna bentoska.[4]
Molekularne i morfološke studije dijele Haptophyta u pet redova a grupiranih u dva razreda:[9]
U filogenetskom stablu, pronađene su sljedeće veze:[10]
Haptophyta CoccolithophoridaeCocolito fosil Discoaster surculus (Coccolithales)
Cvjetanje Emiliania huxleyi (Isochrysidales)
Cvjetanje Phaeocystis (Phaeocystales)
Haptofite ili haptofiti (latinski: Haptophyta ili Prymnesiophyta) su skupina, koljeno jednoćelijskih algi s oko 500 vrsta, uglavnom morskih, za koje je karakteristično prisustvo dva biča, haptoneme (kontraktilni dodatak koji površno nalikuje bičevima) i golih ćelija ili prekrivenih karbonatnim pločama. Uključuje poznate kokolitoforide koje imaju egzoskelet od krečnjačkih plakove ili kokolita, zvanih kokosfera i čine najbogatiju morsku površinu fitoplanktona, posebno u otvorenom moru, a izuzetno su obilne i kao mikrofosili. Neki haptofiti periodično proizvode cvjetanje algi otrovne morske vode ili stvaraju neugodnu pjenu koja se često nakuplja na plažama.
Prymnesiophyceae je razred protista koljena Haptophyta. To su jednoćelijski organizmi sa hloroplast ima, pa se smatraju algama. Neke vrste u životnom ciklusu imaju dvije faze, izmjenjujući kolonijalni ili vlaknasti stadij s drugim bičastim. Razred uključuje većinu vrsta haptofita, uključujući poznate rodove Phaeocystis, Chrysochromulina, Prymnesium i Coccolithophore, koji povremeno izazivaju cvjetanje algi na obalama i otvorenoj pučini, sa vrlo vidljivim utjecajem na funkcioniranje morskih ekosistema.[1]
Prymnesiophyceae je razred protista koljena Haptophyta. To su jednoćelijski organizmi sa hloroplast ima, pa se smatraju algama. Neke vrste u životnom ciklusu imaju dvije faze, izmjenjujući kolonijalni ili vlaknasti stadij s drugim bičastim. Razred uključuje većinu vrsta haptofita, uključujući poznate rodove Phaeocystis, Chrysochromulina, Prymnesium i Coccolithophore, koji povremeno izazivaju cvjetanje algi na obalama i otvorenoj pučini, sa vrlo vidljivim utjecajem na funkcioniranje morskih ekosistema.
Τα απτόφυτα ή απτοφύκη, γνωστά και ως πρυμνεσιόφυτα, απαντώνται σε μεγάλη ποικιλία και αριθμό στις θάλασσες. Τα περισσότερα απτόφυτα είναι μονοκύτταρα και πολλά είδη είναι μαστιγοφόρα, ενώ άλλα στερούνται μαστιγίου ή δημιουργούν νηματώδη στάδια. Ωστόσο μερικά μπορεί να έχουν ακόμα αμοιβαδοειδή ή κοκκώδη στάδια. Διαδραματίζουν σημαντικό ρόλο στην παγκόσμια βιογεωχημεία και στα τροφικά πλέγματα των φυσικών οικοσυστημάτων και των συστημάτων υδατοκαλλιεργειών.
Τα απτόφυτα έλαβαν την ονομασία τους από την παρουσία ενός χαρακτηριστικού μαστιγίου, γνωστό ως απτόνημα. Μερικές φορές ωστόσο χρησιμοποιείται και ο όρος Haptophyceae. Πρόσφατες μοριακές φυλογενετικές αναλύσεις, οι οποίες γενικώς υποστηρίζουν προηγούμενες ταξινομήσεις που βασίζονται σε μορφολογικά γνωρίσματα όπως το απτόνημα, επιβεβαιώνουν ότι τα απτόφυτα είναι μια μονοφυλετική ομάδα. Παρ’ όλα αυτά, αν και η φυλογενετική ταξινόμηση χρησιμοποιείται όλο και περισσότερο τα τελευταία χρόνια, υπάρχουν κάποιες αμφιβολίες σχετικά με το ποιο ταξινομικό επίπεδο είναι καταλληλότερο γι’ αυτά.
Τα κύτταρα των απτοφύτων περιέχουν μιτοχόνδρια που εμφανίζουν σωληνοειδείς ακρολοφίες και συχνά διαθέτουν ένα ή δύο κίτρινα ή χρυσοκαστανά πλαστίδια εγκλεισμένα σε ενδοπλασματικό δίκτυο. Συγκεκριμένα τα πλαστίδια αυτά είναι συνήθως χλωροπλάστες που επειδή περιέχουν και πρόσθετες φωτοσυνθετικές χρωστικές, καλύπτεται το πράσινο των χλωροφυλλών (χλωροφύλλη α, c1,2) εμφανίζοντας διαφορετικό χρώμα. Τα πλαστίδια των απτοφύτων συχνά έχουν μορφή πεταλούδας (ψυχόμορφα) όταν παρατηρούνται με μικροσκοπία επιφθορισμού. Η επιφάνεια του κυττάρου είναι καλυμμένη με λεπτά λέπια ή κόκκους οργανικής σύνθεσης. Επιπλέον μερικές φορές υπάρχουν και ασβεστοποιημένα λέπια (κοκκόλιθοι) που είναι ορατά με το οπτικό μικροσκόπιο. Τα είδη που φέρουν κοκκόλιθους ονομάζονται αντίστοιχα κοκκολιθοφόρα. Οι κοκκόλιθοι συντίθενται συνήθως από κρυστάλλους ανθρακικού ασβεστίου υπό μορφή ασβεστίτη και δεν αποδομούνται εύκολα στις φυσιολογικές τιμές pH των ωκεανών. Συνεπώς παρατηρείται αυξημένη συσσώρευσή τους στον πυθμένα τους. Επιπλέον τα κύτταρα των απτοφύτων εμφανίζουν κατά κανόνα δύο μαστίγια ή κανένα, παρά το γεγονός ότι μερικά είδη του γένους Chrysochromulina διαθέτουν τέσσερα μαστίγια. Τα δύο μαστίγια ίσως έχουν όμοια δράση (ομοδυναμικά) ή κινούνται διαφορετικά (ετεροδυναμικά). Τα μαστιγοφόρα κύτταρα μπορεί να φέρουν επίσης ένα χαρακτηριστικό απτόνημα και ένα σωματικό κάλυμμα από οργανικές ή/και ενασβεστωμένες φολίδες.
Το απτόνημα ξεκινάει από τον κορυφαίο πόλο του κυττάρου, μεταξύ των μαστιγίων. Το στέλεχος του απτονήματος περιλαμβάνει έξι ή επτά μεμονωμένους μικροσωληνίσκους τοποθετημένους υπό την μορφή δακτυλίου ή ημισεληνοειδώς, περιβαλλόμενους από ένα δακτύλιο ενδοπλασματικού δικτύου. Το απτόνημα μπορεί να είναι αρκετά βραχύ ή πολύ μακρύτερο σε σχέση με το σώμα του κυττάρου. Σε αντίθεση όμως με τα μαστίγια, το απτόνημα δεν μπορεί να πάλλεται, αλλά μπορεί να κάμπτεται και συνήθως να περιελίσσεται.
Η περιέλιξη του απτονήματος φαίνεται ότι λειτουργεί ως μέσο αποφυγής εμποδίων. Κατά την επαφή με ένα εμπόδιο, το απτόνημα συσπειρώνεται γρήγορα και τα μαστίγια αλλάζουν την κατεύθυνση του παλμού, προωθώντας το κύτταρο με ταχύτητα προς τα πίσω. Πειράματα έχουν αποκαλύψει ότι η περιέλιξη προκαλείται από ταχεία εισροή Ca2+ στο κύτταρο από το περιβάλλον του. Στις διαδικασίες της θρέψης και της αποφυγής συγκρούσεων, η συμπεριφορά μαστιγίων και απτονήματος φαίνεται να είναι συντονισμένη. Σε ορισμένες περιπτώσεις το απτόνημα βοηθά στην σταθεροποίηση των κυττάρων των απτοφύτων σε υποστρώματα.
Οι πλέον κοινοί τύποι φολίδων που παράγονται από τα απτόφυτα είναι οι λεπτές, ωοειδείς ή κυκλικές οργανικές φολίδες και οι κοκκόλιθοι. Οι οργανικές φολίδες είναι ο αρχαιότερος τύπος φολίδων στα απτόφυτα. Είναι δομημένες σε μεγάλο βαθμό από κυτταρίνη και στο ηλεκτρονικό μικροσκόπιο εμφανίζουν ινώδη μορφή. Η καταγωγή αυτών των φολίδων προέρχεται από τις κιστέρνες ενός μεμονωμένου πολύ μεγάλου σωματίου Golgi, το οποίο βρίσκεται κοντά στα μαστιγιακά βασικά σωμάτια. Αυτή η ασυνήθης συσκευή Golgi είναι χαρακτηριστική των απτοφύτων. Οι ώριμες οργανικές φολίδες συναντώνται στην εξωτερική επιφάνεια των κυττάρων. Τα κοκκολιθοφόρα ωστόσο παράγουν δύο κύριους τύπους κοκκόλιθων. Τους ετεροκοκκόλιθους που σχηματίζονται στο εσωτερικό των κυττάρων και στην συνέχεια εκκρίνονται στην επιφάνεια, αποτελούμενοι από μάλλον λίγους συμπλεκόμενους κρυστάλλους ασβεστίτη και τους ολοκοκκόλιθους. Αυτοί σχηματίζονται εξωτερικά και αποτελούνται από έναν μεγάλο αριθμό μικροσκοπικών κρυστάλλων ασβεστίτη και δεν εμπλέκονται αλλά συγκρατούνται με οργανικό υλικό. Η ενασβέστωση στα απτόφυτα που σχηματίζουν κοκκόλιθους εξαρτάται σε μεγάλο βαθμό από τη φωτοσύνθεση. Η φωτοσύνθεση παρέχει υψηλής ενέργειας μόρια που είναι απαραίτητα για τις διεργασίες μεταφοράς στις κυτταρικές μεμβράνες και στις μεμβράνες Golgi. Η φωτοσύνθεση δρα επίσης και ως παγίδα για το διοξείδιο του άνθρακα κατευθύνοντας έτσι την τελική αντίδραση για την ενασβέστωση. Έχει προταθεί ότι οι κοκκόλιθοι συμμετέχουν σε λειτουργίες όπως η εστίαση του φωτός εντός των κυττάρων, στο περιορισμό της πρόσβασης παθογόνων μικροοργανισμών και ιών στο εσωτερικό των κυττάρων των απτοφύτων όπως και στην παροχή προστασίας έναντι της θήρευσης από μικρά φυτοφάγα.
Οι πληθυσμοί των απτοφύτων αυξάνονται μέσω μιτωτικής κυτταρικής διαίρεσης. Η μίτωση κατά κανόνα συμβαίνει κατά τη διάρκεια της νύχτας, προφανώς επειδή η ολοκλήρωση αυτής της διαδικασίας σε αυτό τον χρόνο έχει ως αποτέλεσμα τη μικρότερη διατάραξη της φωτοσύνθεσης.
Τα περισσότερα απτόφυτα είναι θαλάσσια, εντοπισμένα σε σημαντικούς αριθμούς σε πολικά, υποπολικά, εύκρατα και τροπικά ύδατα ενώ έχει περιγραφεί τουλάχιστον ένα είδος που αναπτύσσεται σε γλυκό νερό. Ένας αριθμός από περίεργες μορφές εντοπίζονται σε σκοτεινούς ωκεανούς βάθους μεγαλύτερου των 200m. Τα απτόφυτα είναι σημαντικά από οικολογική άποψη όσον αφορά στις βιοτικές αλληλεπιδράσεις αλλά και στην βιογεωχημεία. Στις βιοτικές αλληλεπιδράσεις περιλαμβάνονται οι ρόλοι των απτοφύτων ως πηγή τροφής στα φυσικά τροφικά πλέγματα και σε εκείνα των υδατοκαλλιεργειών αλλά και η παραγωγή τοξινών από ορισμένα είδη. Τα απτόφυτα λόγω του μικρού μεγέθους, της ταχείας αύξησής τους, της ευπεψίας και του θρεπτικού περιεχομένου τους θεωρούνται ως υψηλής ποιότητας τροφή για το θαλάσσιο ζωοπλαγκτόν.
Μερικά είδη αρκετών γενών απτοφύτων επηρεάζουν τα βιοτικά συστήματα με την παραγωγή τοξινών ή άλλων επιβλαβών ουσιών. Για παράδειγμα το ευρέως διαδεδομένο είδος μαστιγοφόρου Chrysochromulina polylepis μπορεί να παράγει τοξικές «ανθίσεις» στην ανοικτή θάλασσα που προκαλούν τον θάνατο ψαριών και ασπονδύλων.
Επιπλέον τα κοκκολιθοφόρα επηρεάζουν την παγκόσμια ανακύκλωση άνθρακα μέσω των κοκκολίθων, οι οποίοι συμβάλλουν σημαντικά στην συσσώρευση ανθρακικών στα βαθιά θαλάσσια ύδατα. Τα απτόφυτα που σχηματίζουν «ανθίσεις», είναι επίσης γνωστά για την παραγωγή μεγάλων ποσοτήτων διμεθυλοσουλφιδίου, ενός πτητικού θειούχου μορίου που ενισχύει την όξινη βροχή. Επειδή οι κοκκόλιθοι είναι συνήθως μικροί και παρουσιάζουν χαμηλό ενδημισμό (περιορισμό ορισμένων ειδών σε συγκεκριμένες τοποθεσίες), χρησιμοποιούνται ευρέως ως στρωματογραφικοί δείκτες για την ταύτιση των πετρωμάτων αντίστοιχης ηλικίας από διαφορετικές περιοχές. Οι απολιθωμένοι κοκκόλιθοι χρησιμοποιούνται ευρέως ως βιοδείκτες στη βιομηχανία πετρελαίου και ως δείκτες συνθηκών του παρελθόντος σχετικών με το κλίμα και τη χημεία των ωκεανών.
Μέχρι στιγμής έχουν περιγραφεί έως και 280 είδη απτοφύτων, πολλά βάσει διακριτών γνωρισμάτων των οργανικών ή ανοργανοποιημένων φολίδων τους. Επιπλέον η μοριακή εκτίμηση γενών ενδέχεται να αποκαλύψει έναν βαθμό πολυφυλίας που απαιτεί την καθιέρωση νέων γενών.
Τα απτόφυτα ή απτοφύκη, γνωστά και ως πρυμνεσιόφυτα, απαντώνται σε μεγάλη ποικιλία και αριθμό στις θάλασσες. Τα περισσότερα απτόφυτα είναι μονοκύτταρα και πολλά είδη είναι μαστιγοφόρα, ενώ άλλα στερούνται μαστιγίου ή δημιουργούν νηματώδη στάδια. Ωστόσο μερικά μπορεί να έχουν ακόμα αμοιβαδοειδή ή κοκκώδη στάδια. Διαδραματίζουν σημαντικό ρόλο στην παγκόσμια βιογεωχημεία και στα τροφικά πλέγματα των φυσικών οικοσυστημάτων και των συστημάτων υδατοκαλλιεργειών.
The haptophytes, classified either as the Haptophyta, Haptophytina or Prymnesiophyta (named for Prymnesium), are a clade of algae.
The names Haptophyceae or Prymnesiophyceae are sometimes used instead.[2][3][4] This ending implies classification at the class rank rather than as a division. Although the phylogenetics of this group has become much better understood in recent years, there remains some dispute over which rank is most appropriate.

The chloroplasts are pigmented similarly to those of the heterokonts,[5] but the structure of the rest of the cell is different, so it may be that they are a separate line whose chloroplasts are derived from similar red algal endosymbionts.
The cells typically have two slightly unequal flagella, both of which are smooth, and a unique organelle called a haptonema, which is superficially similar to a flagellum but differs in the arrangement of microtubules and in its use. The name comes from the Greek hapsis, touch, and nema, round thread. The mitochondria have tubular cristae.
The best-known haptophytes are coccolithophores, which make up 673 of the 762 described haptophyte species,[6] and have an exoskeleton of calcareous plates called coccoliths. Coccolithophores are some of the most abundant marine phytoplankton, especially in the open ocean, and are extremely abundant as microfossils, forming chalk deposits. Other planktonic haptophytes of note include Chrysochromulina and Prymnesium, which periodically form toxic marine algal blooms, and Phaeocystis, blooms of which can produce unpleasant foam which often accumulates on beaches.[7]
Haptophytes are economically important, as species such as Pavlova lutheri and Isochrysis sp. are widely used in the aquaculture industry to feed oyster and shrimp larvae. They contain a large amount of polyunsaturated fatty acids such as docosahexaenoic acid (DHA), stearidonic acid and alpha-linolenic acid.[8] Tisochrysis lutea contains betain lipids and phospholipids.[9]
The haptophytes were first placed in the class Chrysophyceae (golden algae), but ultrastructural data have provided evidence to classify them separately.[10] Both molecular and morphological evidence supports their division into five orders; coccolithophores make up the Isochrysidales and Coccolithales. Very small (2-3μm) uncultured pico-prymnesiophytes are ecologically important.[7]
Haptophytes was discussed to be closely related to cryptomonads.[11]
Haptophytes are closely related to the SAR clade.[12]
Subphylum Haptophytina Cavalier-Smith 2015 [Haptophyta Hibberd 1976 sensu Ruggerio et al. 2015][13]
The haptophytes, classified either as the Haptophyta, Haptophytina or Prymnesiophyta (named for Prymnesium), are a clade of algae.
The names Haptophyceae or Prymnesiophyceae are sometimes used instead. This ending implies classification at the class rank rather than as a division. Although the phylogenetics of this group has become much better understood in recent years, there remains some dispute over which rank is most appropriate.
Las haptofitas (Haptophyta o Prymnesiophyta) son un grupo de algas unicelulares de unas 500 especies, fundamentalmente marinas, que se caracterizan por la presencia de dos flagelos, un haptonema (apéndice contráctil superficialmente parecido a un flagelo) y células desnudas o cubiertas de placas calcáreas.[3][4][5] Incluye a los conocidos cocolitóforos, que tienen un exoesqueleto de placas calcáreas o cocolitos denominado cocósfera y constituyen el fitoplancton marino más abundante, especialmente en mar abierto, además de ser extremadamente abundantes como microfósiles. Algunas haptofitas periódicamente producen floraciones algales marinas tóxicas o generan una espuma desagradable que a menudo se acumula en las playas.
Las haptofitas son un componente importante del fitoplancton marino y debido a su alta concentración juegan un papel importante como productores primarios en estos ecosistemas. En aguas tropicales son más abundantes, constituyendo alrededor del 45% del fitoplancton en las latitudes medias del Atlántico Sur. Disminuyen en abundancia hacia las latitudes templadas, aunque algunas haptofitas todavía se encuentran en aguas polares. Unas pocas especies viven en agua dulce o en el suelo. Muchas especies son simbiontes de otros organismos, por ejemplo, forman asociaciones con radiolarios.
Algunas especies periódicamente producen floraciones algales marinas tóxicas o generan una espuma desagradable que a menudo se acumula en las playas. Algunas especies segregan potentes exotoxinas que causan un gran mortalidad entre peces y moluscos.
Las haptofitas son unicelulares, aunque a veces se agrupan en colonias o filamentos. El ciclo de vida generalmente comprende una sola etapa, pero en algunos casos alterna entre etapas flageladas y no flageladas. La etapa flagelada es unicelular y planctónica, mientras que la no flagelada suele ser colonial y bentónica. Las formas bentónicas pueden ser filamentosas, cocoides o palmeloides.[6]
Las haptofitas son organismos principalmente fotosintéticos, si bien algunos miembros son mixótrofos o heterótrofos. Muchas especies son fagotrofas y presentan vesículas en el citoplasma con las que digieren el material ingerido. No son selectivas en cuanto al material alimenticio que introducen en las vesículas y este puede consistir tanto de desechos orgánicos como de bacterias y otros pequeños organismos.[7]
Las células de las haptofitas tienen típicamente dos flagelos iguales o levemente desiguales, y lisos. La excepción es Pavlovales, donde uno de los flagelos es más largo que el otro y están cubiertos de pequeños pelos o escamas. Las haptofitas pueden nadar indistintamente con los flagelos dirigidos hacia delante o hacia la parte posterior del cuerpo. El movimiento es rápido y a menudo realizan cambios de dirección.
A menudo presentan un único orgánulo denominado haptonema, que es superficialmente similar a un flagelo pero que se diferencia del mismo en el conjunto de microtúbulos y en el uso. Su nombre proviene de las raíces griegas hapsis (tacto) y nema (hilo). Dependiendo de la especie, el haptonema puede ser largo o corto, y estar enrollado en espiral o no. Se encuentra situado entre los dos flagelos y no bate como estos, pero puede encogerse y estirarse y su función sería la fijación al sustrato, intervenir en el movimiento o apoyar la capacidad fagotrófica de algunas haptofitas. El haptonema está constituido por siete microtúbulos simples rodeados por la membrana del retículo endoplasmático.
Aunque se incluyen algunas formas con células desnudas (Pavlovales), la mayoría presentan células cubiertas de placas o escamas, de forma típicamente elíptica. Las placas orgánicas se originan en el aparato de Golgi y están a menudo envueltas en mucílago. Las placas pueden estar sin mineralizadar, consistiendo principalmente en carbohidratos, pero también pueden estar calcificadas, siendo muy raras las placas de sílice.
Las especies que tienen las placas calcificadas se conocen con el nombre de cocolitóforos. Estos tienen un exoesqueleto de placas calcáreas denominado cocolito y constituyen el fitoplancton marino más abundante, especialmente en mar abierto. La arquitectura de los cocolitos es muy variada, a menudo con una compleja ornamentación de espinas y rebordes, y se conocen especies con forma elíptica, pentágono, estrella, disco, etc. Los cocolitóforos se incluyen en los órdenes Coccolithales e Isochrysidales y son extremadamente abundantes como microfósiles. Estos se conocen desde el Carbonífero, hace unos 300 millones de años, y son muy abundantes durante el Jurásico.
Por lo general, las haptofitas presentan en cada célula de uno a cuatro cloroplastos discoidales alargados con un pirenoide en el centro del cloroplasto o como un abultamiento a un lado. Los tilacoides están agrupados de tres en tres y carecen de lamela periférica. Los cloroplastos están rodeados por cuatro membranas, estando la más externa cubierta de ribosomas y conectada con la membrana nuclear, formando el retículo endoplasmático cloroplástico. Como pigmentos fotosintéticos presentan clorofilas a, c1, c2 y c3 (esta última en Prymnesiophyceae). Como pigmentos accesorios presentan xantofilas de tipo dianinoxantina, diatoxantina y fucoxantina, además de β-caroteno. Como sustancias de reserva utilizan crisolaminarina, paramilo y gotas de lípidos, que se forman en vacuolas fuera del cloroplasto.
Se considera que los cloroplastos de las haptofitas proceden de la endosimbiosis secundaria de un alga roja. Los cloroplastos se pigmentan de forma similar a Ochrophyta (heterocontofitas, como en las algas doradas), pero se diferencian en el resto de la estructura celular, así que es posible que sean una línea evolutiva separada en la que los cloroplastos se deriven de endosimbiosis independientes.[8]
De la superficie de la célula se pueden originar seudópodos y filipodios, que pueden ser lineales o ramificados. Muchos son fagotrofos y en consecuencia tienen vacuolas alimenticias en el citoplasma con las que digieren bacterias y otros pequeños organismos. Presentan también un gran aparato de Golgi situado en el extremo anterior de la célula y, en algunos casos, también una vacuola contráctil. Algunos pocos géneros presentan una mancha ocular sensible a la luz, por ejemplo, Pavlova y Diacronema.[9] La mancha ocular se sitúa dentro o próximo al cloroplasto y consiste en gotitas de lípidos.[10] Las mitocondrias tienen crestas tubulares.
Las haptofitas usualmente se producen asexualmente por bipartición y la mitosis es abierta. La reproducción sexual puede ser común en algunos grupos.[2] Se ha encontrado algunas especies con un ciclo diplohaplonte, en la cual una etapa diploide flagelada alterna con una etapa haploide filamentosa. La etapa diploide es planctónica mientras que la haploide es bentónica.[6]
Ciertas especies como Emiliania huxleyi producen floraciones algales y algunos géneros como Chrysochromulina y Prymnesium producen toxinas que son fatales para los peces. Phaeocystis produce floraciones que generan una espuma desagradable que a menudo se acumula en las playas. Algunas haptofitas como Pavlova lutheri e Isochrysis son económicamente importantes, pues se utilizan extensivamente en la acuicultura.
Estudios moleculares y morfológicos dividen a Haptophyta en cinco órdenes agrupados en dos clases:[2]
Se han encontrado las siguientes relaciones:[11]
Haptophyta CoccolithophoridaeLas algas haptofitas se habrían originado por simbiogénesis entre una célula huésped hetrótrofa haptista y un alga endosimbionte heterokonta de acuerdo con la genética plastidial;[12] este endosimbionte sería una ocrofita de tipo Diatomista por la presencia de clorofila c3. Es por esto que haptofitas y ocrofitas tienen muchas similitudes, pues comparten la presencia de las clorofilas a, c1, c2, c3, β-caroteno y xantofilas como diatoxantina y fucoxantina, además de tener afinidades en la estructura de los tilacoides.[13]
Cocolito fósil de Discoaster surculus (Coccolithales)
Floración de Emiliania huxleyi (Isochrysidales)
Floración de Phaeocystis (Phaeocystales)
Las haptofitas (Haptophyta o Prymnesiophyta) son un grupo de algas unicelulares de unas 500 especies, fundamentalmente marinas, que se caracterizan por la presencia de dos flagelos, un haptonema (apéndice contráctil superficialmente parecido a un flagelo) y células desnudas o cubiertas de placas calcáreas. Incluye a los conocidos cocolitóforos, que tienen un exoesqueleto de placas calcáreas o cocolitos denominado cocósfera y constituyen el fitoplancton marino más abundante, especialmente en mar abierto, además de ser extremadamente abundantes como microfósiles. Algunas haptofitas periódicamente producen floraciones algales marinas tóxicas o generan una espuma desagradable que a menudo se acumula en las playas.
Haptofüüdid (Haptophyta) ehk prümnesiofüüdid on kromalveolaatide hõimkond. Enamik haptofüüte elab meres, vähesed ka magevees.
Haptofüütidel on olemas kloroplastid ja nad on valdavalt planktilised. Neil on kaks viburit ja sünapomorfne haptoteema. Haptofüütide suurus jääb 2–20 mikromeetri vahele.
Haptofüüdid (Haptophyta) ehk prümnesiofüüdid on kromalveolaatide hõimkond. Enamik haptofüüte elab meres, vähesed ka magevees.
Haptofüütidel on olemas kloroplastid ja nad on valdavalt planktilised. Neil on kaks viburit ja sünapomorfne haptoteema. Haptofüütide suurus jääb 2–20 mikromeetri vahele.
Tarttumalevät (Haptophyta) on yksisoluisia levälajeja käsittävä Chromalveolata-kunnan kaari. Valtaosa tunnetuista lajeista elää meressä. Lajeja on kuvattu noin 500.
Varhaisimmat tarttumalevien fossiilit on löydetty kivihiilikaudelta 300 miljoonan vuoden takaa.
Tarttumaleväsolussa on kaksi uintisiimaa solun etupäässä tai kyljessä. Monilla lajeilla on lisäksi uintisiimojen välissä kierteinen siima eli haptoneema, jonka avulla solut voivat tilapäisesti kiinnittyä alustaansa. Tarttumalevien nimitys tulee tästä rakenteesta. Soluissa on kaksi kahden kalvon ympäröimää viherhiukkasta, joissa on a- ja c-klorofylliä sekä beetakaroteenia ja fukoksantiinia. Jälkimmäisin pigmentti antaa leville ruskehtavan värin. Yhteyttämispigmentit muistuttavat ruskeiden levien pigmenttejä. Chromalveolata-kunnalle tyypillisesti tarttumalevien mitokondrioiden sisäkalvojen poimut eli kristat ovat putkimaiset.
Solua peittää kerros selluloosasta koostuvia suomuja. Joillain lajeilla (lahko Coccolithales) solun pinnalla on kalkkipitoisia suomuja eli kokkoliitteja. Liitukaudella 65-95 miljoonaa vuotta sitten näitä tarttumaleviä esiintyi runsaasti, ja niiden sedimentoituneista kalkkikuorista muodostui suuria liitukivikerrostumia. Näistä kerrostumista liitukausi on saanut nimensä.
Tarttumalevät (Haptophyta) on yksisoluisia levälajeja käsittävä Chromalveolata-kunnan kaari. Valtaosa tunnetuista lajeista elää meressä. Lajeja on kuvattu noin 500.
Varhaisimmat tarttumalevien fossiilit on löydetty kivihiilikaudelta 300 miljoonan vuoden takaa.
Tarttumaleväsolussa on kaksi uintisiimaa solun etupäässä tai kyljessä. Monilla lajeilla on lisäksi uintisiimojen välissä kierteinen siima eli haptoneema, jonka avulla solut voivat tilapäisesti kiinnittyä alustaansa. Tarttumalevien nimitys tulee tästä rakenteesta. Soluissa on kaksi kahden kalvon ympäröimää viherhiukkasta, joissa on a- ja c-klorofylliä sekä beetakaroteenia ja fukoksantiinia. Jälkimmäisin pigmentti antaa leville ruskehtavan värin. Yhteyttämispigmentit muistuttavat ruskeiden levien pigmenttejä. Chromalveolata-kunnalle tyypillisesti tarttumalevien mitokondrioiden sisäkalvojen poimut eli kristat ovat putkimaiset.
Solua peittää kerros selluloosasta koostuvia suomuja. Joillain lajeilla (lahko Coccolithales) solun pinnalla on kalkkipitoisia suomuja eli kokkoliitteja. Liitukaudella 65-95 miljoonaa vuotta sitten näitä tarttumaleviä esiintyi runsaasti, ja niiden sedimentoituneista kalkkikuorista muodostui suuria liitukivikerrostumia. Näistä kerrostumista liitukausi on saanut nimensä.
Les Haptophytes, ou Haptophyta, sont une division d'algues unicellulaires (du groupe des Chromalveolata) — elles sont parfois appelées Prymnesiophyta, mais ce nom n'a jamais été validé — caractérisées par la présence d'un appendice particulier différent des flagelles, l'haptonème (appendice filiforme contenant des microtubules, dont la taille varie selon l’espèce, et qui permettrait l’adhésion à un substrat, le déplacement de particules voire la capture de proies).
Le nombre d'espèces actuelles est estimé à environ 500. De nombreuses espèces sont tropicales, quelques espèces vivent en eau douce et de nombreux groupes fossiles existent.
Ce groupe a présenté (fossiles) et présente encore une grande variété de formes de vie, montrant différents types structuraux.
On distingue des formes monadoïdes, cellules isolées mobiles et flagellées possédant deux flagelles, égaux ou inégaux, avec l'haptonème inséré entre eux ; des formes palmelloïdes, colonies immobiles protégées par un mucilage qui les recouvre ; des formes « coccoïdes », cellules seules immobiles ; voire des formes amoeboïdes caractérisées par des cellules mobiles à pseudopodes.
Au niveau ultra-structural, un réticulum endoplasmique entoure la cellule sous la membrane plasmique et un autre réticulum enveloppe le noyau et les plastes. Dans ces derniers, les thylakoïdes sont assemblés par trois, sans lamelle périphérique, ne formant pas de grana. Les pigments principaux sont les chlorophylles a et c accompagnées aussi de xanthophylles, comme la fucoxanthine. De plus les mitochondries ont des crêtes tubulaires.
Ces caractères rapprochent les Haptophyta des Heterokontophyta. Ceux-ci présentent en plus une hélice de transition à l'insertion flagellaire, une lamelle périphérique de thylakoïdes, et des mastigonèmes (poils tubulaires) sur les flagelles.
En phylogénétique, un caractère synapomorphique, ou synapomorphie, est un caractère dérivé (ou apomorphique), partagé par deux ou plusieurs taxons-frères. Pour les Haptophyta cette synapomorphie se manisteste par les caractères suivants :
Les cellules ont deux flagelles typiques de longueur inégale, tous les deux lisses, et un organite unique, l’haptonème, qui ressemble à un flagelle de l’extérieur, mais a un arrangement différent des microtubules à l’intérieur et a un usage différent. Le nom vient du grec hapsos, toucher, et nema, fil. Les mitochondries ont des crêtes tubulaires.
La plupart des Haptophytes sont les Coccolithophoridés (ou Prymnesiophyceae), qui sont ornées d’écailles calcifiées appelées coccolithes, et sont parfois trouvés dans les microfossiles. Parmi les autres Haptophytes planctoniques notables, les espèces des genres Chrysochromulina et Prymnesium sont périodiquement responsables d’efflorescences algales marines toxiques. Les données moléculaires et morphologiques soutiennent toutes les deux la division des Haptophytes en cinq ordres ; les Coccolithophoridés rassemblant les Isochrysidales et les Coccolithales.
La division des Haptophyta est composée de deux classes bien distinctes au niveau ultra-structural : les Pavlovophyceae et les Prymnesiophyceae. Cette scission dans le groupe a longtemps été suggérée par de nombreuses études ultrastructurales, notamment par rapport à l'insertion flagellaire. Plus récemment[Quand ?], la comparaison de la sous-unité 18S a renforcé cette séparation.
En 2021, Kawachi et al. y ont ajouté une troisième classe.
Selon AlgaeBase (13 jan. 2022)[4] :
Les Haptophytes, ou Haptophyta, sont une division d'algues unicellulaires (du groupe des Chromalveolata) — elles sont parfois appelées Prymnesiophyta, mais ce nom n'a jamais été validé — caractérisées par la présence d'un appendice particulier différent des flagelles, l'haptonème (appendice filiforme contenant des microtubules, dont la taille varie selon l’espèce, et qui permettrait l’adhésion à un substrat, le déplacement de particules voire la capture de proies).
Le nombre d'espèces actuelles est estimé à environ 500. De nombreuses espèces sont tropicales, quelques espèces vivent en eau douce et de nombreux groupes fossiles existent.
Ce groupe a présenté (fossiles) et présente encore une grande variété de formes de vie, montrant différents types structuraux.
On distingue des formes monadoïdes, cellules isolées mobiles et flagellées possédant deux flagelles, égaux ou inégaux, avec l'haptonème inséré entre eux ; des formes palmelloïdes, colonies immobiles protégées par un mucilage qui les recouvre ; des formes « coccoïdes », cellules seules immobiles ; voire des formes amoeboïdes caractérisées par des cellules mobiles à pseudopodes.
Au niveau ultra-structural, un réticulum endoplasmique entoure la cellule sous la membrane plasmique et un autre réticulum enveloppe le noyau et les plastes. Dans ces derniers, les thylakoïdes sont assemblés par trois, sans lamelle périphérique, ne formant pas de grana. Les pigments principaux sont les chlorophylles a et c accompagnées aussi de xanthophylles, comme la fucoxanthine. De plus les mitochondries ont des crêtes tubulaires.
Ces caractères rapprochent les Haptophyta des Heterokontophyta. Ceux-ci présentent en plus une hélice de transition à l'insertion flagellaire, une lamelle périphérique de thylakoïdes, et des mastigonèmes (poils tubulaires) sur les flagelles.
Haptophyta, diklasifikasikan baik sebagai Prymnesiophyta (dinamai untuk Prymnesium) atau Haptophyta, adalah sebuah divisi dari ganggang.
Nama-nama Haptophyceae atau Prymnesiophyceae kadang-kadang digunakan sebagai gantinya.[1][2][3] Akhiran ini menyiratkan klasifikasi di peringkat kelas bukan sebagai divisi. Meskipun filogenetik dari kelompok ini telah menjadi jauh lebih baik dipahami dalam beberapa tahun terakhir, masih ada beberapa sengketa tentang peringkat yang paling tepat.
Haptophyta, diklasifikasikan baik sebagai Prymnesiophyta (dinamai untuk Prymnesium) atau Haptophyta, adalah sebuah divisi dari ganggang.
Nama-nama Haptophyceae atau Prymnesiophyceae kadang-kadang digunakan sebagai gantinya. Akhiran ini menyiratkan klasifikasi di peringkat kelas bukan sebagai divisi. Meskipun filogenetik dari kelompok ini telah menjadi jauh lebih baik dipahami dalam beberapa tahun terakhir, masih ada beberapa sengketa tentang peringkat yang paling tepat.
Haptophyta (ook wel Prymnesiophyta genoemd) zijn fotoautotrofe algen. Ze vormen een stam behorend tot het rijk Chromalveolata. Haptophya zijn samen met de diatomeeën de belangrijkste groep van fytoplankton in de huidige oceanen.
De gele, geelbruine of bruine chloroplasten bevatten chlorofyl a en c, β-caroteen en xanthofyl. Als reservestof worden chrysolaminarine, olie en soms ook paramylon (paramylum) opgeslagen. De chloroplasten bevinden zich in een plooi van het endoplasmatisch reticulum. De chloroplasten lijken op de andere heterokonten, de celopbouw wijkt echter af, waardoor de Haptophyta tegenwoordig ook systematisch van de heterokonten afgesplitst zijn.
Zweepcellen hebben twee meestal even lange of bijna even lange zweepstaartjes. De zweepstaartjes zijn niet zoals bij de overige heterokonten met dunne vederharen bezet, maar met submicroscopisch kleine schubben of knobbels. In tegenstelling tot de heterokonten hebben de zweepstaartjes ook geen opgezwollen basis.
Naast de zweepstaartjes hebben de cellen ook een haptonema. Dat is een draadvormig aanhangsel waarmee de cel zich kan vastzetten. De bouw lijkt oppervlakkig gezien op een zweepstaartje, maar op doorsnee zijn 6-7 in een sikkelvorm liggende microtubuli te zien.
De cellen hebben aan de buitenkant schubben bestaande uit polysachariden (meest cellulose), die in de golgiblaasjes gevormd en aan de buitenkant van de cel afgezet worden. Bij de Coccolithophorales zijn de buitenste schubben verkalkt en worden coccolieten genoemd. De functie van de coccolieten wordt tot nu toe nog niet erg begrepen. Vermoed wordt dat ze onder andere dienen ter bescherming tegen vijanden, opstijgvermogenregulering, lichttoevoer of verkalking ter ondersteuning van de fotosynthese. Ook kan een oogvlek zoals bij de goudwieren (Chrysophyta) en de Euglenozoa aanwezig zijn.
Hoewel de vertegenwoordigers van de monadalen het meest voorkomen, zijn er ook capsale, kokkale en trichale vertegenwoordigers. Sommige soorten hebben een heteromorfe generatiewisseling, dat wil zeggen dat de diploïde en de haploïde generatie uiterlijk goed onderscheidbaar zijn. Bij deze kalkalgen wisselt de diploïde generatie, waarbij de algen zweepstaartjes bezitten en als plankton leven met een haploïde generatie, waarbij ze benthisch zijn, dat wil zeggen dat ze dan op de zeebodem leven.
Slechts weinig soorten leven in zoetwater, de meeste leven als plankton in de zee. Met een grootte van minder dan 20 µm worden ze tot het kalkige nanoplankton gerekend. Ze kunnen in zeer grote hoeveelheden optreden en een overwegend deel van het zeeplankton uitmaken. Daarmee spelen de planktonische Haptophyten in de zee een belangrijke rol als primaire producent. Enkele soorten komen over de hele wereld verspreid voor, de meeste soorten komen echter in de gematigde streken voor. In het bijzonder komt in grote hoeveelheden de wereldwijd verbreide soort Emiliania huxleyi voor. Zij behoort tot de belangrijkste producenten van biologisch calciumcarbonaat en kan een grote algenbloei geven. Andere planktonische haptophytes, zoals Chrysochromulina en Prymnesium zorgen periodiek voor een giftige algenbloei, terwijl Phaeocystis bij algenbloei een onplezierig schuim vormt dat zich dikwijls ophoopt aan de kusten.
De coccolieten van de kalkalgen vormden in het verre verleden via sedimentering en diagenese kalkgesteente. Coccolieten zijn onder anderen een belangrijk bestanddeel van de krijtrotsen van Rügen, Møn en de Zuid-Engelse krijtkust van Dover. Eén cm³ krijt bestaat uit rond de 800 miljoen coccolieten. De vroegste vondsten van de Coccolithoforen stammen uit het Trias. De grootste verbreiding en vormenrijkdom vindt tijdens het Krijt plaats, waarbij aan het eind van dit tijdperk massale sterfte optrad. Een nieuw hoogtepunt werd ongeveer 50 miljoen jaar geleden in het Eoceen bereikt. Vertegenwoordigers van de familie Braarudosphaera kwamen al in het krijt voor.
Aan de hand van fossielresten van kalkalgen in de sedimenten kan de wetenschap de ouderdom van deze sedimenten en de toen heersende klimaatomstandigheden vaststellen.
Kalkalgen spelen in de natuur een zeer belangrijke rol:
Kalkalgen zijn in de zee de op afstand productiefste kalkvormers. Daarmee staan ze aan de oorsprong van talrijke gesteenten (vooral kalksteen en dolomiet). Ze beïnvloeden ook de chemische samenstelling van het zeewater en daarmee hangt de opnamecapaciteit van het broeikasgas koolstofdioxide hoofdzakelijk af van hun biologische en kalkbindende activiteit.
Tot de Haptophyta behoren:
Ook wordt er wel maar één klasse onderscheiden: de Haptophyceae met ongeveer 500 soorten en 75 geslachten in 4 orden.
Haptophyta (ook wel Prymnesiophyta genoemd) zijn fotoautotrofe algen. Ze vormen een stam behorend tot het rijk Chromalveolata. Haptophya zijn samen met de diatomeeën de belangrijkste groep van fytoplankton in de huidige oceanen.
De gele, geelbruine of bruine chloroplasten bevatten chlorofyl a en c, β-caroteen en xanthofyl. Als reservestof worden chrysolaminarine, olie en soms ook paramylon (paramylum) opgeslagen. De chloroplasten bevinden zich in een plooi van het endoplasmatisch reticulum. De chloroplasten lijken op de andere heterokonten, de celopbouw wijkt echter af, waardoor de Haptophyta tegenwoordig ook systematisch van de heterokonten afgesplitst zijn.
Zweepcellen hebben twee meestal even lange of bijna even lange zweepstaartjes. De zweepstaartjes zijn niet zoals bij de overige heterokonten met dunne vederharen bezet, maar met submicroscopisch kleine schubben of knobbels. In tegenstelling tot de heterokonten hebben de zweepstaartjes ook geen opgezwollen basis.
Naast de zweepstaartjes hebben de cellen ook een haptonema. Dat is een draadvormig aanhangsel waarmee de cel zich kan vastzetten. De bouw lijkt oppervlakkig gezien op een zweepstaartje, maar op doorsnee zijn 6-7 in een sikkelvorm liggende microtubuli te zien.
De cellen hebben aan de buitenkant schubben bestaande uit polysachariden (meest cellulose), die in de golgiblaasjes gevormd en aan de buitenkant van de cel afgezet worden. Bij de Coccolithophorales zijn de buitenste schubben verkalkt en worden coccolieten genoemd. De functie van de coccolieten wordt tot nu toe nog niet erg begrepen. Vermoed wordt dat ze onder andere dienen ter bescherming tegen vijanden, opstijgvermogenregulering, lichttoevoer of verkalking ter ondersteuning van de fotosynthese. Ook kan een oogvlek zoals bij de goudwieren (Chrysophyta) en de Euglenozoa aanwezig zijn.
Hoewel de vertegenwoordigers van de monadalen het meest voorkomen, zijn er ook capsale, kokkale en trichale vertegenwoordigers. Sommige soorten hebben een heteromorfe generatiewisseling, dat wil zeggen dat de diploïde en de haploïde generatie uiterlijk goed onderscheidbaar zijn. Bij deze kalkalgen wisselt de diploïde generatie, waarbij de algen zweepstaartjes bezitten en als plankton leven met een haploïde generatie, waarbij ze benthisch zijn, dat wil zeggen dat ze dan op de zeebodem leven.
Svepeflagellatene (Haptophyta) er en divisjon av encella alger. Typisk har cellene to flageller av ulik lengde og de har to gyllenbrune kloroplaster
Flere arter skiller ut giftstoffer som kan drepe fisk, f.eks. Chrysochromulina polylepis som ved den store algeoppblomstringen på Sørlandet i 1988 førte til omfattende fiskedød.[1]
Svepeflagellatene (Haptophyta) er en divisjon av encella alger. Typisk har cellene to flageller av ulik lengde og de har to gyllenbrune kloroplaster
Flere arter skiller ut giftstoffer som kan drepe fisk, f.eks. Chrysochromulina polylepis som ved den store algeoppblomstringen på Sørlandet i 1988 førte til omfattende fiskedød.
Haptofity (Haptophyta) – typ (gromada) glonów z supergrupy Chromalveolata (Chromista).
Haptofity to grupa glonów o nie do końca ustalonej pozycji systematycznej. W XX w. organizmy te grupowano jako Coccolithinae wśród złotowiciowców i określano polską nazwą wiciowce wapienne[1]. Na początku XXI w. większość badaczy umieszczała je wśród chromistów (bądź chromalweolatów) jako odrębną linię ewolucyjną niezwiązaną ani ze stramenopilami, ani z alweolatami. W podobnej sytuacji znajdują się kryptomonady i czasem te dwie linie są łączone jako Hacrobia[2]. Jedne badania z XXI w. potwierdzają bliskość haptofitów i kryptomonad[3][4], podczas gdy inne wskazują na ich dużą odrębność, potwierdzając bliskość haptofitów i pozostałych chromalweolatów (oraz ameb Rhizaria), a zbliżając kryptomonady do linii rozwojowej obejmującej rośliny[5].
W jednym z systemów zaproponowanych przez Cavaliera-Smitha haptofity mają rangę infrakrólestwa Haptista, które jest monotypowe, zawierając jedynie typ Haptophyta. Jest on tam równoważny infrakrólestwu Heterokonta, z którym tworzy podkrólestwo Chromobiota, które z kolei z podkrólestwem Cryptista (zawierającym szeroko ujmowane kryptomonady) tworzy królestwo Chromista[6].
Od drugiej połowy XX w. haptofity dzieli się na dwie główne grupy:
W rejestrze AlgaeBase do drugiej połowy 2012 r. zgromadzono informacje o 385 gatunkach Coccolithophyceae, 15 gatunkach Pavlovophyceae i 125 gatunkach skupionych w kilku rodzinach nieprzypisanych do żadnej z tych grup[7]. Za najbardziej liczebnego przedstawiciela haptofitów kokolitotwórczych uważana jest Emiliania huxleyi[8].
Haptofity mają postać wiciowców, często, ale nie zawsze ruchliwych – jednokomórkowych monad lub ich kolonii, np. w formie nitkowatej lub palmelloidalnej, przy czym u niektórych gatunków stadium kolonijne występuje na przemian z monadalnym. Zdarzają się też postacie pełzakowate. Pod względem rozmiaru niektóre sytuują się wśród organizmów nanoplanktonowych, podczas gdy inne tworzą widoczne gołym okiem kolonie (Phaeocystis). Charakterystycznym, niewystępującym poza tą grupą organellum jest haptonema, czyli wyrostek przypominający wić. Może on przybierać różne postacie i długości (u Pavlovophyceae i niektórych Coccolithophyce raczej krótka, a u Isochrysidales może wręcz być jej brak). Wici u Coccolithophyceae równe lub jedna nieco dłuższa od drugiej, zwykle dwie, umiejscowione mniej lub bardziej szczytowo. Natomiast u Pavlovophyceae jedna wić jest zdecydowanie krótsza od drugiej, a położone są po przeciwnych końcach komórki. Przednia wić Pavlovophyceae pokryta jest wyrostkami. U Coccolithophyce wici mogą zawierać substancję autofluoroscencyjną, czego nie ma u Pavlovophyceae. Z kolei u tych ostatnich mogą występować niezwiązane z wiciami plamki oczne[9]. Retikulum endoplazmatyczne położone pod samą błoną komórkową, wnikając do haptonemy, jednak pozostawiając lukę u nasady wici. Aparat Golgiego położony blisko kinetosomów wici i haptonemy. Jego cysterny leżą prostopadle do dłuższej osi komórki, układając się wachlarzowo. Aparat Golgiego u gatunków wytwarzających pancerzyki jest zaangażowany w to działanie. Liczba chloroplastów od 1 do 4 (u niektórych zanikły)[9], choć podawany jest też zakres 1–2[10]. W chloroplastach znajduje się pirenoid. Zewnętrzna błona chloroplastów przechodzi w zewnętrzną błonę jądra. Barwniki fotosyntetyczne to chlorofile a, c1, c2 i c3 (ostatni u Coccolithophyceae), β-karoten, diadinoksantyna i diatoksantyna. Głównym produktem zapasowym często jest chryzolaminaryna[9][10]. Chloroplasty haptofitów powstały na drodze wtórnej endosymbiozy, pochodząc od krasnorostów. Są spokrewnione z chloroplastami kryptomonad i stramenopili[8]. Realizowane strategie pokarmowe to autotrofizm, miksotrofizm i heterotrofizm[9][10].
Wielu przedstawicieli podklasy Coccolithophyce wytwarza organiczne płytki, wśród nich zaś grupa określana jako kokolitofory, pokrywa je wapiennymi skorupkami zwanymi kokolitami (stąd nazwa)[9][10]. Na podstawie badań filogenetycznych sugeruje się, że jest to historycznie cecha powszechna w tej podklasie, a niezdolność do tworzenia kokolitów jest zjawiskiem wtórnym[11]. Płytki pancerzyka haptofitów mają różną budowę, a często ten sam gatunek wytwarza różne ich typy w zależności od fazy ploidii. Za najpierwotniejszą cechę uważa się brak jakichkolwiek płytek, typowy dla podklasy Pavlovoohyceae. Niektóre Coccolithophyceae (np. Chrysochromulina) wytwarzają jedynie płytki zbudowane z substancji organicznej, bez kokolitów. Inne wytwarzają kokolity w fazie diploidalnej, a w fazie haploidalnej jedynie płyki organiczne. Nie jest jasne, czy jest to cecha pierwotna ewolucyjnie czy też wtórna – rozważane są też obie opcje, np. według jednej z hipotez Emiliania zachowała ją po swoich przodkach, podczas gdy przodkowie Pleurochrysis wytwarzali kokolity w obu fazach, a następnie utracili tę zdolność. Dość często występuje model, w którym w fazie haploidalnej wytwarzane są inne rodzaje kokolitów (nanolity, holokokolity) niż w fazie diploidalnej (heterokokolity). Typowe kokolity są zbudowane z kalcytu, podczas gdy nanolity mogą być aragonitowe[12].
Wśród haptofitów zdarzają się gatunki słodkowodne, jednak zdecydowana większość żyje w wodach słonych, zarówno przybrzeżnych, jak i pełnomorskich[10]. Haptofity nierzadko osiągają bardzo duże zagęszczenia, zajmując istotną pozycję w fitoplanktonie niemal wszystkich środowisk morskich. Czasem powodują zakwity wód. Ze względu na tworzenie kokolitów odgrywają znaczącą rolę w obiegu węgla i wapnia, a ich szczątki od późnego triasu są ważnym elementem wapiennych osadów. Pośrednio, przez wiązanie dwutlenku węgla, ma to wpływ na kształtowanie klimatu. Rozległe zakwity są widoczne na zdjęciach satelitarnych. Mają one wpływ na biocenozy oceaniczne, jako że mogą być toksyczne dla innych organizmów[13]. Pomory ryb wywołane zakwitami wód w słonawych duńskich jeziorach powodowanymi przez Prymnesium parvum opisywano już w pierwszej połowie XX w[14], natomiast jest możliwe, że podobne oddziaływanie na ekosystem od czasu do czasu haptofity wywierały przez ok. 200 milionów lat[13]. Rodzaj Phaeocystis jest kosmopolityczny, a w chłodniejszych rejonach oceanu może dominować w fitoplanktonie[15].
Wpływ haptofitów na klimat, zwłaszcza wytwarzających kokolity, jest złożony. Kokolity otaczające komórki odbijają światło w charakterystyczny sposób, co w przypadku zakwitu daje wrażenie białej wody. Ma to pewien wpływ na albedo obszaru objętego zakwitem. Według modeli rozpraszania światła słonecznego docierającego do oceanu wynika, że istnienie warstwy zajętej przez kokolitofory zwiększa albedo oceanu, sprawiając że najpłytsza jego warstwa (kilka metrów) jest jaśniejsza, a głębsza ciemniejsza niż wynikałoby to z samego pochłaniania przez glony. Może to prowadzić do redukcji strefy eufotycznej, preferowanej przez organizmy fotosyntetyzujące, z ponad 24 m do 8 m w strefie zakwitu. Bardziej naświetlona warstwa przypowierzchniowa ogrzewa się, a mniej naświetlona ochładza, co powoduje powstanie stratyfikacji termicznej i zmniejsza mieszanie się wody. Jednocześnie strefa rozjaśniona przez światło odbijane od kokolitów zwiększa strefę fotoinhibicji. Sam więc wpływ kokolitów (nawet bez wpływu toksycznego innych substancji wytwarzanych przez haptofity) ostatecznie może zmniejszyć produkcję pierwotną strefy objętej zakwitem. Jednocześnie jednak ze względu na skalę przestrzenną i czasową takich zakwitów, ich wpływ na całkowite albedo oceanu uważany jest za nieistotny[16]. Do wytwarzania wapiennych struktur zużywany jest dwutlenek węgla. W związku z tym organizmy je produkujące w pewnych warunkach (np. dostępności substancji odżywczych) mogą zwiększać tę produkcję przy wzroście stężenia CO2. W przypadku organizmów wodnych jednak może zachodzić przeciwny proces, tj. rozpuszczanie struktur wapiennych przez CO2 powodujący zakwaszanie wód. Ze względu na specyficzny sposób wytwarzania kokolitów, zakwaszenie ma słabszy wpływ, przez co w ciągu XIX i XX w. średnia masa kokolitów wzrosła o 40%, a eksperymenty w warunkach laboratoryjnych sugerują, że kokolitofory mogą rozwijać się lepiej przy nieco podwyższonym stężeniu CO2[17]. Badania w różnych warunkach jednak dają zróżnicowane wyniki. Kokolitofory zasadniczo dostosowują się do różnych warunków, o czym świadczy stan kokolitów zdeponowanych w czasach zlodowaceń, gdy stężenie dwutlenku węgla było niskie[18].
Haptofity (Haptophyta) – typ (gromada) glonów z supergrupy Chromalveolata (Chromista).
Haptophyta ou Prymnesiophyta, também chamadas de "algas douradas/amarronzadas" por conta da presença de pigmentos acessórios, é um grupo de algas unicelulares ou coloniais. Com cerca de 300 espécies distribuídas em, aproximadamente, 80 gêneros[1], as haptófitas constituem o fitoplâncton e são essencialmente marinhas (embora também possa se encontrar formas de água doce e raras formas terrestres). Os seus cloroplastos pigmentam-se de forma similar aos Heterokontophyta (como nas algas douradas, mas diferenciam-se no resto da estrutura celular, pelo que podem ser considerados uma linha evolutiva separada, na qual os cloroplastos são derivados de endossimbiontes similares. As mitocôndrias possuem cristas tubulares.
Quando monadais, as células possuem a presença de dois flagelos iguais (podendo, também, haver a ocorrência de flagelos desiguais, como no caso do gênero Pavlova) que são lisos, e uma estrutura estritamente característica do grupo denominada haptonema, que é superficialmente similar a um flagelo mas que se diferencia do mesmo pelo conjunto de microtúbulos e em sua função. O filo pode receber duas nomeações distintas entre os pesquisadores: a primeira é Haptophyta, que vem do grego, hapsis, tacto e de nema, hilo, evidenciando a presença marcante do haptonema; ou Prymnesiophyta, por conta do gênero Prymnesium, importante devido à ocorrência de florações.
Os Haptophyta mais conhecidos são os cocolitóforos (ordens Coccolithales e Isochrysidales), que possuem em sua superfície celular a presença de placas/escamas orgânicas que sofrem um processo de calcificação denominadas cocolitos (o conjunto de cocolitos é denominado cocolitosfera)[2]. Constituem o fitoplâcton marinho mais abundante, especialmente em mar aberto e é extremamente abundante como microfóssil. Outros Haptophyta plactónicos incluem Chrysochromulina e Prymnesium, que periodicamente produzem blooms de algas tóxicos, e Phaeocystis que produz blooms que geram uma espuma desagradável que se acumula nas praias. Estudos moleculares e morfológicos dividem Haptophyta em cinco ordens. Um exemplo de cocolitoforídeo é a Emiliania huxleyi que pode formar florações cobrindo milhares de quilômetros quadrados no oceano.
Haptophyta ou Prymnesiophyta, também chamadas de "algas douradas/amarronzadas" por conta da presença de pigmentos acessórios, é um grupo de algas unicelulares ou coloniais. Com cerca de 300 espécies distribuídas em, aproximadamente, 80 gêneros, as haptófitas constituem o fitoplâncton e são essencialmente marinhas (embora também possa se encontrar formas de água doce e raras formas terrestres). Os seus cloroplastos pigmentam-se de forma similar aos Heterokontophyta (como nas algas douradas, mas diferenciam-se no resto da estrutura celular, pelo que podem ser considerados uma linha evolutiva separada, na qual os cloroplastos são derivados de endossimbiontes similares. As mitocôndrias possuem cristas tubulares.
Quando monadais, as células possuem a presença de dois flagelos iguais (podendo, também, haver a ocorrência de flagelos desiguais, como no caso do gênero Pavlova) que são lisos, e uma estrutura estritamente característica do grupo denominada haptonema, que é superficialmente similar a um flagelo mas que se diferencia do mesmo pelo conjunto de microtúbulos e em sua função. O filo pode receber duas nomeações distintas entre os pesquisadores: a primeira é Haptophyta, que vem do grego, hapsis, tacto e de nema, hilo, evidenciando a presença marcante do haptonema; ou Prymnesiophyta, por conta do gênero Prymnesium, importante devido à ocorrência de florações.
Os Haptophyta mais conhecidos são os cocolitóforos (ordens Coccolithales e Isochrysidales), que possuem em sua superfície celular a presença de placas/escamas orgânicas que sofrem um processo de calcificação denominadas cocolitos (o conjunto de cocolitos é denominado cocolitosfera). Constituem o fitoplâcton marinho mais abundante, especialmente em mar aberto e é extremamente abundante como microfóssil. Outros Haptophyta plactónicos incluem Chrysochromulina e Prymnesium, que periodicamente produzem blooms de algas tóxicos, e Phaeocystis que produz blooms que geram uma espuma desagradável que se acumula nas praias. Estudos moleculares e morfológicos dividem Haptophyta em cinco ordens. Um exemplo de cocolitoforídeo é a Emiliania huxleyi que pode formar florações cobrindo milhares de quilômetros quadrados no oceano.
Fästalger (Haptophyta) är en division marina och akvatiska alger. Totalt finns omkring 300 arter, de flesta i saltvatten.[1] På svenska kallas gruppen omväxlande för fästalger[2][3] eller haptofyter. Karakteristiskt för gruppen är haptonemat, ett slags piskliknande (fäst)organ samt att många arter är förkalkade (dessa benämns kockolitoforer). Fästalgerna är viktiga både för kolets och svavlets kretslopp. De kan också orsaka omfattande algblomningar. Bergarten krita består av sedimenterade kockolitoforer.[2]
Under första hälften av 1900-talet räknades fästalgerna till klassen Chrysophyceae. Det unika haptonemat, uppbyggnad av Golgiiapparaten samt andra ultrastrukturella drag ledde fykologerna till att på 1960-talet tilldela haptofyterna sin egen klass, Haptophyceae. 1976 delade Hibberd i sin tur upp gruppen i olika klasser samlade under sin egen division, Haptophyta. Idag finns två klasser: Prymnnesiophyceae och Pavlovophyceae. [4][2]
Många DNA-analyser (till exempel på rRNA-gener) visar att fästalgerna, haptofyterna, hör hemma inom supergruppen Chromalveolata, [4] till vilken gruppen numera vanligen räknas. Ett antal ultrastrukturella drag och biokemiska parametrar (till exempel bildningen beta-1,3-glukan utanför kloroplasterna, klorofyll a, c, kloroplasternas tylakoidstruktur, fyra membran runt kloroplasten) delas av fästalgerna och heterokonterna.[4][5]
Andra molekylärbiologiska studier visar istället att fästalgerna är släkt med rödalger och gröna växter.[6]
Många studier visar emellertid ett nära släktskap mellan fästalgerna och rekylalgerna.[4][6][5][7] Dessa två grupper räknas tillsammans med några smågrupper in i supergruppen Hacrobia.[7]
Vissa bentiska arter har cellvägg. Kalkfjäll (kockoliter) förekommer hos kockolitoforerna. Haptofyterna har två flageller och beroende på rörelsemönster kan algerna simma såväl framåt som bakåt.[4] Flagellerna är ibland isomorfa, ibland heteromorfa.[1] Haptonemat är en unik cellstruktur som endast finns hos fästalgerna.[8] Det används som fästorgan och kan rullas ihop likt en spiral. Hos kockolitoforerna används haptonemat förmodligen till största delen som ett slags känselorgan för identifiera hinder. När ett hinder påträffas börjar algen simma åt motsatt håll.[8] Några haptofyter saknar eller har endast rester av haptonema och/eller flageller.[1]
Klorofyllet i kloroplasterna innehåller klorofyll a och c, betakaroten, diadinoxantin och diatoxantin. Även olika varianter av fukoxantin förekommer.[1] Vissa arter av Prymnesium har ibland kiselkorn uppbyggda av små plattliknande fjäll.[1] Fästalger i klassen Pavlovophyceae har en ögonfläck som gör att algen kan bestämma ljusets riktning.[4]
Fästalgerna är mycket viktiga primärproducenter som finns i nästan all världens hav. Efter sin död spelar de kalcifierade arterna (kockolitoforerna) en viktig roll för uppkomsten av sedimentära bergarter, närmare bestämt kalkstenen krita. Fästalgerna är genom sitt upptag av kol och inlagring i kalkskalen betydelsefulla för kolets storskaliga kretslopp. Dessutom har fästalgerna stor inverkan på svavlets kretslopp då döda fästalger kan avge svavelrika föreningar, till exempel dimetylsulfoxid (DMSO)[2] och dimetylsulfoniopropionat (DMSP).[9] Vissa föreningar kan brytas ned till dimetylsulfid som sedan kan hamna i atmosfären och oxideras till svaveldioxid, sulfat och svavelsyra. En del av föreningarna stimulerar molnbildningen och påverkar därmed jordens klimat.[1][2][8]
Vissa fästalger, till exempel Phaeocystis, är välkända för att kunna orsaka omfattande algblomningar. Vissa algblomningar är så omfattande att de kan ses på satellitbilder.[1]. 2002 förstördes 90% av växt- och djurarterna i Phan Ri-bukten i Vietnam av en blomning av haptofyten Phaeocystis globosa.[10] Framförallt två släkten, Prymnesium och Chrysochromulina är välkända för att producera toxiner. En art ur det sistnämnda släktet har också orsakat illaluktande algblomningar i en kanadensisk sjö.[1].
Vissa fästalger är inte fotosyntetiserande autotrofer utan lever som heterotrofer, d.v.s. måste ta upp organiskt kol från omgivningen. Andra arter kan ta upp organiskt kol trots egen fotosyntes. Dessa arter har således mixotrofi.[1]
Fästalger (Haptophyta) är en division marina och akvatiska alger. Totalt finns omkring 300 arter, de flesta i saltvatten. På svenska kallas gruppen omväxlande för fästalger eller haptofyter. Karakteristiskt för gruppen är haptonemat, ett slags piskliknande (fäst)organ samt att många arter är förkalkade (dessa benämns kockolitoforer). Fästalgerna är viktiga både för kolets och svavlets kretslopp. De kan också orsaka omfattande algblomningar. Bergarten krita består av sedimenterade kockolitoforer.
Гаптофитовые водоросли, Гаптофиты, Примнезиофитовые водоросли, или Примнезиофиты (лат. Haptophyta) — тип эукариот неясного систематического положения. Включает два класса — Павловофициевые (Pavlovophyceae) и Примнезиофициевые (Prymnesiophyceae).
Гаптофиты — это группа аутотрофных, осмотрофных или фаготрофных простейших, которые населяют морские экосистемы. Гаптофиты, как правило, одноклеточные, но встречаются и колониальные формы. Несмотря на маленький размер, эти организмы играют очень большую и важную роль в геохимических циклах углерода и серы.
Гаптофитовые водоросли, Гаптофиты, Примнезиофитовые водоросли, или Примнезиофиты (лат. Haptophyta) — тип эукариот неясного систематического положения. Включает два класса — Павловофициевые (Pavlovophyceae) и Примнезиофициевые (Prymnesiophyceae).
Гаптофиты — это группа аутотрофных, осмотрофных или фаготрофных простейших, которые населяют морские экосистемы. Гаптофиты, как правило, одноклеточные, но встречаются и колониальные формы. Несмотря на маленький размер, эти организмы играют очень большую и важную роль в геохимических циклах углерода и серы.
定鞭藻门(学名:Haptophyta)是一类真核生物,属于多貌生物定鞭隐藻类,曾被归类于囊泡藻界。1955年Parke等引入了"定鞭"(Haptonema)来描述细胞顶端的第3根鞭毛。其主要特征还有细胞表面包裹鳞片。
定鞭藻门(学名:Haptophyta)是一类真核生物,属于多貌生物定鞭隐藻类,曾被归类于囊泡藻界。1955年Parke等引入了"定鞭"(Haptonema)来描述细胞顶端的第3根鞭毛。其主要特征还有细胞表面包裹鳞片。

ハプト藻類は真核微細藻類の一群である。細胞表面に炭酸カルシウムの鱗片である円石を持つグループは円石藻として有名である。
ハプト藻は細胞直径5〜50μm程度の植物プランクトンで、光合成を行う独立栄養生物である。多くは海洋に生息するが、一部の種は淡水や塩湖にも分布する。外洋域におけるバイオマスは大きく、海洋の一次生産者として重要である。北大西洋などの海域では、ハプト藻が大発生してブルームを形成する事もある。現生のハプト藻は90属300種とも言われるが、その分類は後述する生活環の問題を抱えており、正確な属数・種数は不明である。
ハプト藻に関する最古の記載はエーレンベルク(1836)によるものである。彼はバルト海周辺の石灰岩層から微細な円板状の構造物(円石=coccolith)を発見した。しかし彼は、この構造物を生物由来ではなく、化学的、無機的要因によって生成したものと考えた。その後ハクスリー(1858)が同様の構造物を海底の堆積物の中から発見したが、やはり円石は非生物起源であると考えられた。
円石を初めて生物起源であるとしたのは ウォーリッチ(1860)と ソービー(1861)である。彼らは円石が多数結合して中空の球を形成したものを発見し、coccosphere と命名した。現在この語は、円石を持つ細胞全体を、原形質を含めて表す単語として用いられている。しかしながら彼は円石藻という微細藻の存在を提唱したのではなく、coccosphere を有孔虫の生活環の一部と考えるに留まった。1870年代に入ると再び エーレンベルク の円石非生物由来説が支持されるようになった。特に円石の幾何学的な形状から、炭酸カルシウムの凝結、結晶化によると考えられる事が多かった。
円石の持ち主を微細藻であると提唱したのは ワイヴィル・トムソン(1874)である。この時初めて円石は単細胞藻の外被であると考えられた。その後、coccosphere の中に色素体があるという報告や、Murray とBlackman(1898)による細胞分裂の描写が為されるに至り、単細胞藻としての円石藻-ハプト藻が認識される事となった。分類上のハプト藻は、体制と光合成色素の類似から、古くは不等毛植物門黄金色藻綱に含められていた経緯がある。ハプト植物門として独立したのは近年(1962)である。
ハプト藻の葉緑体は紅藻由来で、光合成色素としてクロロフィルa/c、その他補助色素として種々のカロテノイドを持つ。通常、細胞内に葉緑体は二つあり、四重膜に囲まれている。最外膜は核膜と連絡する。三重チラコイド及び埋没型のピレノイドを持つ。ヌクレオモルフやガードルラメラは存在しない。
ハプト藻は細胞の表面に有機質の鱗片や円石を持つ。珪酸質の鱗片を持つ種も報告されている。
ハプトネマ(ハプト鞭毛)は鞭毛に似た器官で、ハプト藻の細胞に1本だけ備わる。特にChrysochromulina 属で発達している。ハプトネマは細胞膜、及び周縁小胞体(peripheral ER)よりなる3重膜に囲まれ、中は単体の微小管(多くの場合6-9本)で構成される。鞭毛とは異なり微小管はペアを作らず、いわゆる9+2構造ではない。基部は鞭毛の基底小体と隣接し、鞭毛根と共に鞭毛装置を構成する。ハプトネマは鞭毛運動は行わないが屈曲が可能であり、瞬間的にハプトネマを巻き縮めるコイリングと呼ばれる現象や、基物に先端を付着させて滑走するグライディング、餌粒子の収集と食胞への運搬など、多彩な働きをもつ。
一部のハプト藻では、核相の違いによって異なった細胞形態を示すことが報告されている。特に円石藻においては、単相(n)の世代と複相(2n)の世代とで異なった円石を付ける例が頻繁にあり、既に別個に命名され記載された属や種が多い。これは、近年になってハプト藻の培養技術が発達し、その生活環が明らかになるにつれ浮上した問題である。原則として学名は一つの生物に対して一つしか認められない為、各世代が別の学名を持つ現状は憂慮すべき事態である。今後こうした命名の重複が明らかになるにつれ、属名あるいは種名の統廃合が進むと予想される。
착편모조류(着鞭毛藻類, haptophytes)는 프림네시움문(Prymnesiophyta) 또는 착면모조문(Haptophyta)으로 분류되는 조류의 생물 문(門)이다. 착편모조강(Haptophyceae)이라는 용어로도 사용된다.[1][2]
2016년 실라(Silar)에 의한 하크로비아의 계통 분류는 다음과 같다.[3][4]
하크로비아 넓은 의미의 크립티스타 코르비헬리아 엔도헬레아상강 코르비스토마상강 좁은 의미의 크립티스타 팔피토아문 롤로모나스아문 레우크립토스상강 은편모조류 합티스타 육질태양충류 착편모조아문 RappemonadaRappephyceae
Haptomonada






















.png)

.png)
