Type locality: Basil Minns Blue Hole, Great Exuma Island, Exuma Cays, The Bahamas (23º 28’ N, 75º 45’ W); collected in 33-43 m depth of anchihaline cave.
Material examined: Holotype (9 mm, 22 trunk segments; ZMA Rem.204.579) and 1 paratype (13 mm, 24 trunk segments; JvdH 12-01 A1); collected by B. Kakuk and T. Iliffe, 12 Jan. 2003. The holotype is dissected and preserved in alcohol. The paratype is prepared for SEM investigations and will be retained in the research collection of the first author (JvdH).
Etymology. – The specific epithet parabenjamini, meaning ‘nearby benjamini’, alludes to the morphological resemblance of this new species to Speleonectes benjamini.
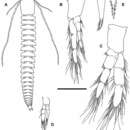
Diagnosis. – A relatively small species (Fig. 4A); pleural tergites well developed, broadly rounded, becoming increasingly pointed in posterior part of trunk; trunk sternites with pointed posterolateral corners; sternal bars heteromorphic; frontal filaments with short, stout processes; antennules with long dorsal flagella; segment 4 of maxillule subrectangular, bearing 1 very long, serrate spine and several slender setae; maxillulary claw very long; maxilla and maxilliped slender, without distinct tagmosis, setation sparse; arc of horseshoe-type claw composed of 7-8 small denticles; anal somite longer than wide; caudal rami shorter than anal somite.
Description. – Body short, up to 13 mm, composed of 24 postcephalic segments (Fig. 4A). Pleural tergites well developed, with broadly rounded distolateral corners on trunk segments 1-7, becoming increasingly acuminate posteriorly. Posterolateral corners of trunk sternites with distinctly pointed corners (Fig. 8B; Pl. 2C). Sternal bars heteromorphic, slightly concave on segments 1-13, posterior to segment 14 as small, convex flaps; segment 14 with large convex flap (Pl. 2C). Male gonopores on trunk segment 14 with rounded lobes.
Cephalon subrectangular, with concave anterolateral excavations, as long as trunk segments 1-4 (Fig. 4A). Frontal filaments bearing short, corpulent, mid-medial processes (Fig. 5A).
Antennules with long dorsal flagella, reaching up to 60% of length of body (Figs. 4A, 5B). Peduncle composed of at least 2 fused segments; ventral margin expanded, bearing densely inserted, long aesthetascs. Dorsal flagellum with 14 segments. Ventral flagellum with up to 15 segments, less than half as long as dorsal flagellum, but slightly longer than head shield.
Antennae (Fig. 5C): Protopod 2-segmented, proximal segment equipped with 7 marginal setae; distal segment with 11 setae. Exopod longer and wider than adjacent distal segment of protopod, bearing about 37 long setae. Endopod forming a semicircular arc; first two proximal segments with 6-8 setae, respectively; distal segment with up to 17 setae arranged in two rows. All setae faintly feathered (cf. Fig. 1E).
Labrum fleshy, with clusters of fine setules on posterior margin (Fig. 6A).
Mandibles (Fig. 6B, C; Pl. 2A): Right incisor process and lacinia mobilis with three large denticles; respectively. Left incisor process with four large denticles; left lacinia mobilis crescent-shaped, apical margin serrate. Molar processes prominent; distal surface long, ovoid.
Maxillules a little more robust than maxillae and maxillipeds (Fig. 7A, D-E). First segment equipped with long and narrow endite bearing 1 large, prominent spine and 5-6 small spines (all naked). Endite of segment 2 ovate, spatulate, with 4-5 long setae on distoanterior margin, and 8 short spines and 1 short seta on distoposterior margin (all short spines naked except 1 rasp-like spine at end of spine row; Fig. 7D). Third segment short, with conical endite, bearing 2 long, slender, rasp-like spines, and a few setae. Segment 4 subrectangular, medial margin even, bearing 1 very long, slender, rasp-like spine on proximal corner (Fig. 7E), and 7-9 long and short setae. Segment 5 as long as segment 4, but slightly narrower, with a cluster of distomedial setae. Sixth segment short, equipped with separate rows of long and shorter setae on distal margins. Claw very long.
Maxillae (Fig. 7B): First endite of first segment bearing 1 apical spine and a few setae; endites 2 and 3 with 1 prominent, apical spine, 3 smaller subapical spines, and several setae. Endite of segment 2 equipped with a single, short spine and about 9 short and long setae. Third segment long, with barely expanded, straight medial margin bearing 10-12 long and short setae. Segment 4 shorter than segment 3; distomedial margin with a row of 10-12 setae. Fifth segment half as long as segment 4, bearing about 6 setae on distomedial margin. Segment 6 distinctly longer than fifth segment, with 4 sparsely inserted setae along medial margin, and separate clusters of setae distally. Arc of horseshoe-type claw composed of 7-8 small denticles flanked by 2 stronger, separate denticles (Fig. 7F).
Maxillipeds long, slender, 9-segmented (Fig. 7C); elbow between segments 4 and 5. Segment 1 bearing a few short medial setae; segment 2 equipped with 2 slender medial spines; segment 3 rather long, with a cluster of distomedial long and short setae. Segment 4 very long and narrow, bearing a row of 7-8 midmedial setae. Segments 5-7 gradually decreasing in length; segment 5 a little shorter than segment 4, distal margin expanded, with a cluster of distomedial setae. Segments 6-8 bearing rows of setae on mid- and distomedial margins, respectively. Segment 8 distinctly longer than segment 7, with several separate clusters of setae on distal margins. Claws subequal to those of maxillae (Fig. 7F; Pl. 2B).
Trunk appendages (Fig. 4B-D): Segment 1 of exopod equipped with long setae on lateral margin, and about 3 serrate spines on distolateral corner (Fig. 4C, E); segment 2 with setae on both lateral and medial margins, and 2-4 serrate spines on distolateral corner; segment 3 ovate, bearing long marginal setae. Endopod slightly shorter and narrower than exopod; distribution of feathered setae similar to that of exopod, with the following exceptions: basal segment with 3-4 rasp-like, lateral spines (Fig. 4E); distolateral corner of segment 2 bearing 4 rasp-like spines and 1 strongly serrate spine, distomedial corner with 2 serrate spines; segment 3 with 2 serrate spines on distolateral corner and 1 serrate spine on distomedial corner. Endopods of anterior and posterior trunk limbs comparatively strongly reduced. All setae faintly feathered (cf. Fig. 1E).
Anal segment longer than wide (Fig 8A); length of caudal rami about 66% of length anal somite; caudal rami bearing 1-2 curling setae on midmedial margins up to 6 curling apical setae, respectively.
Remarks. – Speleonectes parabenjamini is morphologically closely allied to S. benjamini Yager, 1987a. Both species share several distinct apomorphies, for example, antennules with long dorsal flagella; maxillules with rather long claws; trunk sternites with pointed posterolateral corners; and heteromorphic sternal bars. However, some of these synapomorphies are also characterized by interspecific differences. Unlike S. benjamini, S. parabenjamini has less sharply pointed sternites, and the shape of the anterior sternal bars is only slightly concave. Furthermore, S. parabenjamini can be easily distinguished from S. benjamini by possessing ventral antennular flagella that lack a fusion of proximal segments; maxillules and maxillae that are more slender (less expanded), with much longer and slender segments proximal to the elbows; an anal segment that is longer than wide; and caudal rami shorter than the anal segment.
From Koenemann, Iliffe and van der Ham 2003
This species was found during a recent diving expedition on Great Exuma Island located in the central Bahamas, along with two other new sympatric species of remipedes, Speleonectes tanumekes and Speleonectes minnsi.All specimens were collected from the same anchihaline cave between 33 and 43 m depth.
The Basil Minns Blue Hole is located southeast of Georgetown on Great Exuma Island in the central Bahamas. A karst window consisting of two sinkhole entrances at the bottom of a saltwater lake provides access to the submerged cave. A 10-12 m wide passage extends to the northeast from the lake for several hundred meters at 50 m depth, before opening into a 50 m wide collapse-floored room. This circular room contains a large breakdown mound in the center with an air dome above, but no apparent opening to the outside. Several large root masses, penetrating through cracks in the ceiling, hang down into the pool. A Hydrolab DataSonde 3 water quality analyzer carried by a diver was used to profile the water column in the terminal breakdown chamber (Fig. 13). Salinity increased from 9.8 ppt at the surface, to 32.7 ppt at 14 m depth and then more gradually to 34.8 ppt at 46 m depth. Temperature increased from 26.7 degrees C at the surface, to a maximum of 29.1 degrees C between 14 and 19 m and then dropped in several steps to 23.8 degrees C at 46 m depth. Dissolved oxygen (DO) concentration dropped rapidly from 1.06 mg/l at the surface to 0.1 mg/l at 2 m depth and stayed below 0.5 mg/l to 20 m where it increased steadily to 3.5 mg/l at 46 m. The pH at the surface was 7.0; it spiked from 6.9 at 2.6 m to 7.4 at 3.2 m and was followed by a general increase to 7.55 at 46 m depth. The low DO layer corresponded to a murky, hydrogen sulfide zone. Most animals, including remipedes, were observed and collected from the clear, marine saline water below the hydrogen sulfide zone at 25 to 40 m depth. Remipedes were observed swimming in the water column and were collected either by hand in individual vials or with a suction bottle.

Fig. 13. Water column profile of salinity, temperature, dissolved oxygen and pH versus depth in the terminal breakdown room of Basil Minns Blue Hole, Great Exuma Island, Bahamas. Data were collected with a Hydrolab DataSonde 3 water quality analyzer carried by a diver.
Other animals present in the terminal breakdown chamber where the remipedes were collected included copepods (three genera of undescribed epacteriscids and many other unknown species; A. Fosshagen, pers. commun.); the halocyprid ostracodes Danielopolina exuma and Deeveya medix; the leptostracan Speonebalia sp.; the peracarid Thetispelecaris remix; the amphipods Bahadzia sp. and Socarnopsis catacumba; a thermosbaenacean, probably Tulumella sp.; a mysid, probably Stygiomysis sp.; and the polynoid polychaete Pelagomacellicephala iliffei. All of these organisms are cave-adapted, stygobiontic taxa characteristic of hydrologically isolated anchihaline caves. Copepod taxonomist Audun Fosshagen of the University of Bergen considered Basil Minns Blue Hole one of the most interesting copepod caves in the Bahamas.
From Koenemann, Iliffe and van der Ham 2003