Comprehensive Description
provided by Smithsonian Contributions to Zoology
Histioteuthis reversa (Verrill, 1880)
Calliteuthis elongata N. Voss and G. Voss, 1962:184, figs. 4a-f, 6a [new synonym, herein].
Previously unreported collections of reversa comprising approximately 257 juveniles, subadults, and adults mainly taken on cruises of the R/V Anton Dohrn (USNM), R/V Professor Shtokman (IOAN), R/V Vityaz (IOAN), and R/V Walther Herwig (USNM), and recently reported collections of reversa and Histioteuthis elongata (N. Voss and G. Voss, 1962) from Canadian waters (ARC; Stephen, 1982) permitted us to clearly determine that elongata represents the mature stage of the female of reversa, as has been long suspected. The new collections also revealed the presence of unusual, maturity-related photophores in the male, as well as new information on distribution and geographic variations that occur in the species.
The following description is based on large juveniles, subadults, and adults, 35–186 mm ML, from over the full range of the species in the Atlantic and from the western Mediterranean. No specimens were available from the eastern Mediterranean.
DESCRIPTION.—Small to moderately large histioteuthids, females known to mature at 114–186 mm GL, males at about 49–99 mm GL/ML; mantle moderately elongate to elongate (mature females); fins medium-sized, length about 35%–50% ML, width about 40%–60% ML (fins proportionally smaller in mature females); head large, wider than mantle (except in mature females), usually with 1 to 2 weakly developed nuchal folds; eyes markedly asymmetrical except in mature females; arms of medium length, about 100%–150% ML (about 50%–120% ML in mature females), approximately subequal; sucker rings of arms I–III usually smooth except distal-most sucker rings often distinctly incised; sucker rings of arms IV with numerous low, blunt teeth on distal or entire margins; in mature males, suckers on basal portions of all arms with swollen collars, ends of arms I modified with coequal-size suckers on enlarged pedestals; inner web between arms I, II, and III vestigial to low (about 5%–8% of length of longest arm); buccal membrane 7-membered, with second supports single to dorsal sides of arms II or bifurcate to ventral sides of arms I and dorsal sides of arms II, fourth supports without secondary connectives; tentacles 100%–200% ML; club with deep, longitudinal cleft on aboral surface of manus; suckers on manus in about 6 diagonal rows, markedly enlarged in median ventral row to 3–4 times diameter of ventral marginals; sucker rings with numerous straight-sided, blunt or pointed teeth on entire margins, teeth often heavier and more widely spaced on proximal margins; rings of enlarged, median suckers sometimes smooth or irregularly incised.
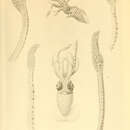
Compound photophores on ventral surface of mantle numerous, intermixed large (arranged in about 7 diagonal rows) and small to near posterior end of mantle; large organs numerous on ventral surface of head; right eye with circlet of 18 organs (17 large and 1 small); basal of arms IV with 4 longitudinal rows of photophores (3 rows of large organs, 1 dorsal marginal row of small organs) (Figure 3e); near-mature and mature females with large, elliptical, black, simple photophores scattered on posterior of ventral surface of elongate mantle and distal of all arms (Figure 3a); mature males with similar but smaller elliptical to round, simple photophores scattered over entire ventral and lateral surfaces and partial dorsal surfaces of mantle, anterior of dorsal surface of head, entire lengths of arms I–III, and entire or distal of arms IV (Figure 3b–d); in males, concentration of organs decreases from arms I to IV.
Dorsal pad of funnel organ with low ridge on median of lateral arms (ridges appear to be independent in small specimens but often appear to merge anteriorly in large individuals); spermatophore short (SpL 2.1–3.6 mm, 2.4%–4.1% ML), sperm mass short to medium (3.7%–25.3% SpL (Atlantic), 30.1%–33.7% SpL (Mediterranean)), cement body medium to long (43.3%–47.4% SpL (Mediterranean), 55.8%–82.5% SpL (Atlantic)), ejaculatory apparatus moderately short (13.2%–19.9% SpL (Atlantic), 20.1%–21.4% SpL (Mediterranean), with single, long loop of inner tube; ejaculatory-apparatus/cement-body connective complex short; mature egg elliptical, average size 1.8 × 1.5 mm; gills about 36%–42% ML (27%–29% ML in mature females), with about 36–42 lamellae in outer demibranch.
Gladius with strong, moderately wide free rachis; vanes diamond-shaped, moderately wide, ending posteriorly in rounded cupped coil; in mature females gladius with short, narrow free rachis and long, narrow, diamond-shaped vanes, widest at anterior – length, with gradual posterior taper ending in narrow cupped coil; lower beak with weak median ridge on lateral wall; ridge line, if projected, would intersect posterior margin of lateral wall at point slightly above free corner; hood somewhat flattened in profile; wing fold high, with steep medial side; radula with first and second laterals asymmetrical and of increasing length, marginal plates weakly developed or absent.
Skin color dark, wine red.
ORIGINAL REFERENCE.—Verrill, 1880:393.
TYPE LOCALITY.—Western Atlantic; 39°53′N, 70°59′W, 666 m.
DEPOSITION OF TYPE.—Holotype: USNM 574849, female, 51 mm ML, R/V Fish Hawk sta 894; fair condition.
DISTRIBUTION.—Confirmed reports show Histioteuthis reversa to be confined to the Atlantic Ocean (Figure 5). The species is widely distributed in the North Atlantic Temperate Region including the eastern and western Mediterranean, and extends northward in the western Atlantic into the subarctic to at least 52°N. The data of Stephen (1982) indicate that reversa is the most common histioteuthid taken off the east coast of Canada. To the south, in a study of the cephalopods taken in Gulf Stream cyclonic rings, Lea (1978) showed a strong association between the occurrence of reversa and temperate slope water. A similar association was found by Lu and Roper (1979) in their investigation of the vertical distribution and abundance of cephalopods at 39°N, 73°W, off the northeastern coast of the United States. Collections taken by the Sargasso Sea Expedition of the R/V Anton Dohrn (USNM), cruise 9 of the R/V Vityaz (IOAN), and cruises of various other vessels show reversa to be widely distributed in the eastern half of the North Subtropical Region and essentially absent in the western half. The species is abundant in the Mauritanean Upwelling and ranges southward in eastern tropical waters to its southern limit at about 23°S. There, in the high productive area off Namibia, reversa has been taken in large numbers by the R/V Akademik Kurchatov and R/V Professor Shtokman (IOAN). The extension of the range of reversa southward in the Benguela Current and into the Agulhas Current by Clarke (1980) on the basis of beaks from stomach contents of sperm whales caught off Donkergat and Durban is questionable. In the northern sector of the Tropical Region, the species extends westward to the northeastern coast of South America (Okutani, 1983) and east of the Lesser Antilles. Histioteuthis reversa is absent from the Caribbean Sea and Gulf of Mexico. It also appears to be absent from the South Subtropical Region. The six individuals reported by Nesis (1974) from the southwestern sector of the region between 35°S and 45°S were examined at the workshop and found to be specimens in poor condition of the closely related atlantica, which is abundant in the waters of the SSTC. A specimen reported by Toll (1982) from the southeastern Pacific was examined earlier (by NV) and found to be atlantica. Retamal and Orellana's (1977) reported capture of reversa from the same area off the southwest coast of South America and Lu and Phillip's report (1985) from the Tasman Sea are probably also cases of misidentification.
The abundance of reversa near slopes of land masses and submarine topography is probably related to the observed preference of this species for waters of higher productivity.
Histioteuthis reversa inhabits the water column from surface waters to in excess of 1000 m. Data from closing nets (Lu and Clarke, 1975a) and open nets (N. Voss, K. Nesis, unpublished data) show small juveniles to about 15 mm ML to be present in the upper 200 m during both day and night. Opening-closing-net captures of all juveniles between depths of 50–630 m during the day and 10–260 m at night (Roper, 1972; Clarke and Lu, 1975; Lu and Clarke, 1975a, 1975b; Lu and Roper, 1979), together with night captures of subadults by open nets fishing as shallow as 80 m, 100 m, and 300 m (N. Voss, K. Nesis, unpublished data) suggest a pattern of diel vertical movement for both growth stages. Daytime, opening-closing-net captures of mature females at 500 m and 1000 m during the day (Lu and Roper, 1979; reported as H. elongata; gladii measurements of two of the three reported specimens corrects the reported sizes of 125 mm ML and 146 mm ML to 180 mm GL and 186 mm GL), daytime, open-net captures of mature females and males in midwaters at 800–1000 m, and nighttime capture in midwaters and near the bottom at approximately 600 m, 640–730 m, 650 m, and 1300 m (N. Voss, K. Nesis, unpublished data) suggest that both sexes mature in deep water. Surface captures of spawning and dead, or near-dead, spent females (Voss and Voss, 1962; Voss, 1969; reported as H. elongata) show that the female of reversa ascends the water column to spawn, and subsequently die, in shallow water.
- bibliographic citation
- Voss, N. A. and Sweeney, M. J. 1998. "Systematics and Biogeography of cephalopods. Volume II." Smithsonian Contributions to Zoology. 277-599. https://doi.org/10.5479/si.00810282.586.277