
Asparagales ye'l nome d'un taxón de plantes perteneciente a la categoría taxonómica d'orde, utilizáu en sistemes de clasificación modernos como'l sistema de clasificación APG III del 2009[3] y l'APWeb,[4] y ta circunscripto obligadamente siquier pola familia Asparagaceae. Asparagales ye un orde namái apocayá reconocíu nos sistemes de clasificación, y munches de les sos families pertenecíen al antiguu orde Liliales. Liliales fuera un orde bien ampliu onde práuticamente s'allugaben toles monocotiledónees de tépalos vistosos y ensin almidón nel endosperma, que fuera malo d'estremar en families porque los sos calteres morfolóxicos nun se daben en patrones que delimitaran grupos claramente. Col tiempu empezar a reconocer l'orde Asparagales, especialmente cuando los analises moleculares d'ADN indicaron que Liliales tenía de ser estremáu siquier en Liliales, Asparagales y Dioscoreales. Asparagales ye un orde abondo grande, consiste en 14 families y unes 26.000 especies. L'orde ta claramente circunscripto sobre la base de los analises de secuencies d'ADN pero ye dificultosu de definir morfolóxicamente, yá que ye unu de los grupos de monocotiledónees estructuralmente más diversu. Asina, l'orde inclúi yerbes de non más de 15 cm hasta delles especies de Agave que superen los 10 m d'altor. Nesti sentíu, la so separación del orde Liliales ye particularmente problemática.
Una de les carauterístiques que definen a esti orde, y posible apomorfía (amás de los datos moleculares), ye la presencia de fitomelanina, un pigmentu escuru presente nes cubiertes seminales. Les fitomelaninas atópase na mayoría de les families de Asparagales, perdiéndose aparentemente en dellos taxones, siendo Orchidaceae l'exemplu más conspicuo nesi aspeutu.
La mayoría de les especies de Asparagales son plantes yerbácees perennes, magar hai dellos exemplos de plantes trepadores (ciertes especies de Asparagus, subfamilia Asparagoideae) y maderices (dellos Aloe, subfamilia Asphodeloideae). Cuasi toles especies d'esti orde producen un recímanu apertáu de fueyes na base de les plantes (son "plantes en roseta"), otres lleven esa mesma disposición de fueyes nel estremu d'un tueru (Yucca), en dellos pocos casos les fueyes producir a lo llargo del tarmu. Les flores frecuentemente tópense na estremidá del tarmu.
Dientro de les monocotiledónees y dende el puntu de vista económicu, l'orde Asparagales ye'l segundu n'importancia dempués del orde Poales (el cual inclúi a los camperes y los ceberaes). L'orde contién munches especies d'importancia hortícola (cebolla, ayu, espárragu, puerru), en floricultura (fresia, gladiolu, iris, munchos miembros d'orquídees), xardinería (Hemerocallis, Convallaria, Agapanthus) y especies (vainilla).
Dende'l puntu de vista de la so taxonomía, Asparagales ye un orde suxuríu por Huber (1977),[5] depués adoptáu por Dahlgren et al. (1985),[6] y reafianzado en 1998 pol Grupu de Filoxenia de les Anxospermes (n'inglés orixinal, "Angiosperm Phylogeny Group", sigles APG),[7] retomáu nel 2003 pol APG II,[8] y nel 2009 pol APG III),[1] y tamién pol APWeb (2001 hasta agora).[4] Les delimitaciones de les families de Asparagales sufrieron una serie de cambeos n'años recién, y entá se precisen más estudios por que se estabilice la so circunscripción y les sos rellaciones.
La mayoría de les especies de Asparagales son plantes yerbácees perennes, magar hai dellos exemplos de plantes trepadores (ciertes especies de Asparagus, subfamilia Asparagoideae de les asparagáceas) y maderices (dellos Aloe, subfamilia Asphodeloideae de les xantorroeáceas). Cuasi toles especies d'esti orde producen un recímanu apertáu de fueyes na base de les plantes (son "plantes en roseta"), otres lleven esa mesma disposición de fueyes nel estremu d'un tueru (Yucca), en dellos pocos casos les fueyes producir a lo llargo del tarmu. Les flores frecuentemente tópense na estremidá del tarmu. Asparagales puede ser estremáu de Liliales polos sos tépalos usualmente non puntiaos, nectarios nos septos de los ovarios (en llugar d'en la base de los tépalos o estambres) y dacuando un crecedera secundaria anómalu (a diferencia de la falta de crecedera secundaria). Los miembros de Asparagales suelen ser geófites, magar nesti casu les sos fueyes son polo xeneral lliniales y ensin venación reticulada fina.
Los sos granes son carauterístiques, cola epidermis esterna de la cubierta seminal obliterada (na mayoría de les especies de frutos carnosos), o presente y con una tiez de fitomelanines negra y carbonácea en munches especies de frutu secu. La parte interna de la cubierta seminal ta usualmente dafechu colapsada. En contraste, les granes de Liliales (morfolóxicamente similares) siempres tienen una epidermis esterna bien desenvuelta, nun tienen fitomelaninas, y usualmente retienen una estructura celular na porción interna de la cubierta seminal.
El caracter inusual d'un crecedera secundaria anómalu con un meristema en fileres superpuestes ("etágeno") ta práuticamente confináu a los xéneros d'esti clado (fora d'esti clado, ta presente namái en Dioscorea). Dalgunes d'estes especies, de los xéneros Agave, Aloe, Cordyline, Dracaena, Nolina, y Yucca, vuélvense árboles curtios y macizos, xeneralmente (pero non siempres) con ramificación llindada (por casu Aloe, Dracaena, Yucca). Otros xéneros como Aphyllanthes y Lomandra, tienen este mesmu tipu de crecedera secundaria pero confináu a los sos tarmos soterraños.
Otros calteres apomórficos del orde son: la presencia d'acedu chelidónico, quiciabes la de velame, quiciabes la de les célules hipodérmiques de los raigaños dimórficas, les anteres más llargues qu'anches, les célules del tapete bi- a tetranucelares, la microsporogénesis simultánea (pero ver n'apartáu de Filoxenia), la presencia d'estilu, les granes con exotesta, el tegmen non persistente, l'endosperma helobial, y la perda del xen mitocondrial sdh3.
Si la microsporogénesis simultánea ye una apomorfía del orde, nel "Nucleu de los Asparagales" aparentemente camudó a socesiva. Hai taxones de clados más basales onde la microsporogénesis ye socesiva, onde los detalles de la formación de la paré son similares a los d'aquellos miembros del nucleu de los Asparagales que fueron estudiaos: son llámines centrífugues ("centrifugal cell plates"). Pero naquellos miembros basales onde la microsporogénesis ye simultánea, la formación de les llámines tamién puede ser centrifuga (Nadot et al. 2006).[9]
Dellos autores consideren al ovariu ínfero una sinapomorfía del orde, pero si dir, foi perdíu tantes vegaes que resulta ser un caracter bien lábil (dientro del orde como nel restu de les anxospermes), polo que la posición del ovariu dientro del orde nun tien valor predictivu.
Tépalos de 3 traces atópase en Orchidaceae, Amaryllidoideae, Iridaceae, Asphodeloideae (pero non en Kniphofia, Asphodelus), Agavoideae, Agapanthoideae, y Hemerocallidoideae; atópense tépalos de 1 traza en Nolinoideae (pero non en Maianthemum stellatum), Allioideae, Aphyllanthoideae, y Asparagoideae. Scilloideae tien tépalos colos dos tipos de vasculatura, inclusive s'atopen tépalos de 5 traces nos verticilos esternos de Urginea.
Paeciera que Asparagales representa una situación dinámica na cual caltiense una variedá de motivos teloméricos en baxa cantidá de copies, en llugar de la repetición dominante de tipu Arabidopsis. Paez qu'en dellos casos, unu de los tipos que taba en baxa cantidá de copies ye amplificáu y reemplaza al tipu estándar. Por casu, saber dende va munchos años que Allium nun tener les repeticiones teloméricas estandarizadas de tipu Arabidopsis, y depués foi afayáu (Adams et al. 2001)[10] que Aloe tampoco les tenía, polo que se predixo que los xéneros entemedios tampoco les tendríen: nun analís de representantes de tolos Asparagales afayóse que, salvu Ornithogalum, nengún tenía les repeticiones de Arabidopsis. Más palantre (Sykorova et al. 2003)[11] afayóse que Ornithogalum y dellos otros xéneros reemplazaren les repeticiones estándar de les plantes por unes de tipu humanu, y que práuticamente tolos xéneros de Asparagales tienen siquier delles copies de les repeticiones teloméricas de tipu humanu, como tamién una variedá d'otres repeticiones allugaes nes terminaciones de les sos cromosomes. La presencia y la composición d'esta variedá de repeticiones teloméricas tienen un gran valor pa discernir el árbol filoxenéticu.
Atopáronse xenomes bien grandes, con un valor C d'unos 350 picogramos, en delles Allioideae, Amaryllidoideae, Scilloideae y Orchidaceae (Leitch et al. 2005).[12]
D'últimes, l'anatomía de la grana paez tener el meyor conxuntu de calteres qu'unifiquen al orde, que d'otra forma ye bien heteroxéneu (particularmente teniendo en cuenta que Orchidaceae ta dientro d'él).
Asparagales comúnmente tien micorrices arbusculares de tipu Arum, ente qu'en Liliales estes micorrizas son comúnmente de tipu Paris (ver F. A. Smith y Smith 1997).[13]
La diversidá taxonómica de les monocotiledónees ta presentada en detalle por Kubitzki (1998,[14] 2006).[15]
De siguío apróvese una llista de la diversidá de Asparagales. Les descripciones son deliberadamente incompletes. Pa más información siga los enllaces.
La familia de les orquídees ye la más grande d'anxospermes. La morfoloxía de les sos flores ye bien distintiva y facer fáciles de reconocer. La flor tien simetría billateral, los 3 sépalos los son coloríos y vistosos (por eso dalgunos llamar tépalos esternos) y hai unu a cada llau y otru nel llau dorsal de la flor. Los 3 pétalos (dacuando llamaos tépalos internos), tamién vistosos, tán allugaos alternadamente ente los sépalos los, dos a los llaos y unu nel llau ventral. Esti postreru llámase "labelo" porque tien una morfoloxía singular. Ente les orquídees atopen dalgunos de los sistemes de polinización más complexos ya interesantes d'ente toles anxospermes.
Les orquídees son conocíes por tener munches especies de gran valor ornamental. La vainilla (de que'l so frutu estrayer la vainillina) tamién ye una orquídea.
Borya ye una planta arborescente y ye típicamente una "planta reviviscente" ("resurrection plant") que s'atopa nes rimaes predreses. Mientres la estación ensuga estes plantes se resecan y vuélvense d'un color anaranxáu óxidu, pero rápido vuélvense verdes y actives una vegada empieza a llover. Xuntu col otru xéneru de boryáceas (Alania) son plantes xeromórficas natives d'Australia, la inflorescencia tien escapo y arreyo, pero les flores son difíciles d'estremar.
Blandfordia ye l'únicu xéneru de la familia Blandfordiaceae, con 4 especies distribuyíes nel este d'Australia. Ellí se les llapada "Christmas Bells" ("campanes de Navidá") por cuenta de la forma de les sos flores y al momentu del so floriamientu, que coincide cola Navidá australiana. Son yerbes perennes erectas (d'unos 1,50 m d'altor), de fueyes dísticas, la so inflorescencia un recímanu, con flores con pedicelos articulaos y tépalos grandes y formando un perigonio tubular. El receptáculu floral desenvuelve un ginóforo. Les anteres son latrorsas y les granes tienen pelos conspicuos.
Lanaria lanata ye la única especie de la familia Lanariaceae, distribuyida nel sur de Sudáfrica. Colos calteres más o menos típicos de les monocotiledónees, pueden ser reconocíes poles sos inflorescencies cortamente ramificaes cubiertes de pelos dendríticos, y los sos flores de simetría radial con tépalos a mediu connar.
Les Asteliáceas formen una familia de 2 a 4 xéneros de plantes presentes nel hemisferiu sur. Son más o menos rizomatusas, de fueyes espiraladas, con flores nuna inflorescencia que puede ser un recímanu o una espiga, les bráctees de les inflorescencies son grandes, les flores pequeñes, los tépalos connados basalmente, el androcéu adnato al perianto, les anteres basifijas.
Les hipoxidáceas entienden unes 150 especies distribuyíes por tol globu salvu Europa y el norte d'Asia. Pueden ser reconocíes polos sos rosetes de fueyes plicadas o siquier plegaes y con bases persistentes, la so indumento non glandular ye abondo prominente. Nes flores, el verticilu esternu de tépalos tiende a ser verde per fora, y l'ovariu ye ínfero, munches vegaes hai una parte delgadamente tubular nel ápice del ovariu formáu polos tépalos connados o per un picu apical del mesmu ovariu.
Esta familia entiende un únicu xéneru, Ixiolirion, con 4 especies distribuyíes dend'Exiptu hasta'l Asia Central. Son yerbes con cormos, les inflorescencies racimosas, y flores curties tubulares azules con ovariu ínfero.
Los 9 xéneros de Tecofileáceas distribuyir en Chile, Estaos Xuníos y África. Son yerbes con cormos, dacuando con fueyes de llámina ancha, peciolaes. Los sos flores tienen tépalos que s'espanden y tienen un androcéu que les vuelve zigomorfas, una y bones los estambres son fuertemente dimórficos. Les anteres abrir por poros.
El xéneru Cyanastrum dacuando ye tratáu na so propia familia Cyanastraceae.
Los dos especies de Doryanthes, l'únicu xéneru de la familia, son yerbes enormes formadores de rosetas, son un elementu conspicuo de la flora de la contorna de Sydney, Australia, y ye difícil qu'el so floriamientu pase inalvertida. Les fueyes tienen márxenes enteros pero se desintegran en fibres escontra'l ápice. Les inflorescencies subumbeladas nacíes a la fin de llargos tarmos tienen numberoses flores de color coloráu brillante, de simetría radial y con ovariu ínfero.
Les iridáceas son unos 70 xéneros con más de 1.600 especies que se distribúin por tol globu.
Estremar d'otres families rellacionaes en que son usualmente yerbes perennes con fueyes xeneralmente ensiformes y unifaciales, la inflorescencia ye una espiga o panícula bracteada de flores solitaries o agrupaciones de visos monocasiales (ripidios), y les flores tienen namái 3 estambres, opuestos a los tépalos esternos.
Dientro d'esta familia atópase Crocus sativus L., de la cual llógrase'l azafrán a partir de los sos estilos secos. Munches especies de Iridaceae presenten una gran importancia económica na horticultura ornamental y na industria de la flor cortada, especialmente Gladiolus, Freesia, Sparaxis, Iris, Tigridia ("flor tigre"), Ixia ("lliriu del maíz"), Romulea, Neomarica, Moraea ("lirio mariposa"), Nemastylis, Belamcanda, Sisyrinchium ("campera de güeyos azules"), Crocosmia, y Trimezia. Munchos otros xéneros (Watsonia, Crocus, Dietes, Tritonia, Hesperantha y Neomarica) cultivar en xardinos en rexones tropicales y templaes, como plantes perennes y bulboses.[16][17][18]
Moraea y Homeria son dos xéneros de plantes venenoses y representen un problema nes rexones productores d'ovinos y bovinos, notablemente en Sudáfrica.[16] Los cormos de delles especies son utilizaos como alimentu por dellos pueblos indíxenes.
Les xeronematáceas, col so únicu xéneru Xeronema, tán distribuyíes na islla Poor Knights de Nueva Zelanda, y en Nueva Caledonia. Son yerbes bastante grandes que pueden ser reconocíes poles sos fueyes isobifaciales equitantes, y les sos inflorescencies atarraquitaes con flores abondo grandes, de simetría radial, que miren escontra riba. Los estambres extán fuertemente exsertos. La familia ye ruinamente conocida entá.
Hemerocallidaceae inclúi plantes yerbácees, glabras, perennes, provistes de curtios rizomes y raigaños fibrosos o rizomatosas con tubérculos radicales. Les fueyes son arrosetadas na base de la planta, alternes, dísticas, planes, sésiles, simples, linear o llanceolaes, paralelinervadas. Los márxenes de la llámina son enteros. Les flores son leve o acusadamente cigomorfas, hermafrodites y presenten nectarios. Tópense dispuestes en dellos tipos d'inflorescencies, tales como visu helicoidal y recímanu, na estremidá d'un escapo bracteado. Inclúi 8 xéneros y aproximao 85 especies distribuyíes en zones templaes d'Europa y Asia, Malasia, India, Madagascar, África y el Pacíficu, especialmente Australia y Nueva Zelandia; hasta Suramérica. Dos de los xéneros, Hemerocallis y Phormium, cultívense como ornamentales en tol mundu.
Les xantorroeoideas tán distribuyíes por Australia. Pueden ser reconocíes pol so vezu, yá que'l so tarmu ye erecto y maderizu (y con un indumento persistente de fueyes seques si nun hai quemes), y nel so estremu tien un agregamientu trupu de fueyes llargues y delgaes. La inflorescencia ye de tipu espiga, erecta y densamente apertada. Les flores son pequeñes y el frutu una cápsula.
Ye una subfamilia de Xanthorrhoeaceae nativa de rexones templaes a tropicales del Vieyu Mundu, con 15 xéneros y 780 especies. La mayor diversidá asocede en Sudáfrica, usualmente en hábitats grebos. Estremar d'otros clados rellacionaos por ser yerbes o árboles paquicaulos (esto ye, con un tueru más anchu nel so parte basal, la parte ancha funcionando n'almacenamientu), con fueyes usualmente ensundioses, flores trímeras con un ovariu súpero, y les granes con arilo. El xéneru más conspicuo de la familia ye Aloe. Munches especies de Aloe son usaes medicinalmente o en cosméticos. Por casu, el "aloin" ye deriváu del Aloe vera y del Aloe ferox, y tien importantes usos médicos, por casu como laxantes y en tratamientos de quemadures, como en productos pa la piel, el pelo y la salú. Miembros de munchos xéneros son usaos como ornamentales, por casu Aloe, Haworthia, Gasteria, Kniphofia, Asphodelus y Bulbine.
Agapanthus ye l'únicu xéneru d'esta subfamilia de les amarilidáceas, reinal de Sudáfrica. Son plantes yerbácees, perennes, más bien robustes, provistes de curtios rizomes y fueyes arrosetadas, linear-oblongas, planes, más bien carnoses. Les flores son polo xeneral grandes, azules o blanques y tópense dispuestes n'umbeles na estremidá d'un escapo áfilo, más llargu que les fueyes. Les inflorescencies topen protexíes por bráctees soldaes ente sigo a lo llargo d'unu de los sos llaos. L'ovariu ye súpero. Nun presenten el golor a ayu carauterísticu de les Allioideae y los analises moleculares d'ADN demuestren que son más cercanes a les Amaryllidoideae. Sicasí, dixébrase d'esti últimu clado pol tipu d'ovariu, la presencia de saponines y l'ausencia de los alcaloides típicos de les amarilidoideas.
Les alioideas pueden atopase xeneralmente nes rexones templaes, templaes y subtropicales de tol mundu. Son herbales y perennes, y de normal tienen bulbu, anque en dellos casos tienen rizoma. Les especies que componen la familia pueden ser reconocíes fácilmente pol so golor carauterísticu (el golor de los ayos y les cebolles, abondo singular como pa ser denomináu golor aliáceo), poles sos fueyes bastante nidies y carnoses y la so inflorescencia asemeyada a una umbela na estremidá d'un escapo, que lleva flores pequeñes a medianes d'ovariu súpero. Cultivar como hortolices y condimenticias, como plantes melecinales y tamién como ornamentales. Nel primer casu, el xéneru económicamente más importante ye Allium, qu'inclúi dalgunes de les plantes comestibles más utilizaes, tales como la cebolla (Allium cepa), l'ayu (A. sativum y tamién A. scordoprasum), la chalota, echalote o escalonia (Allium ascalonicum), el puerru (A. ampeloprasum), el puerru selvaxe o rampla (Allium tricoccum) y dellos condimentos como la cebolleta (A. schoenoprasum). Per otru llau, los compuestos organosulfurados de les aliáceas tienen propiedaes antiosidantes, antibiótiques, anticarcinogénicas, antiteratogénicas y estimulatorias del sistema inmune y protectores de la función hepática.[19]
La familia tamién tien importantes ornamentales.
Esta familia entiende 59 xéneros y aproximao 800 especies mesmes de rexones sobremanera templaes y templaes, de tolos continentes. Son plantes herbales perennes con bulbu, y pueden ser fácilmente reconocíes polos sos fueyes bastante carnoses, les sos flores xeneralmente grandes y curioses, con 6 estambres y un ovariu ínfero, les flores tópense solitaries o, más frecuentemente, dispuestes n'inflorescencies en umbela na estremidá d'un escapo.
Munches de les especies de amarilidoideas son populares como plantes ornamentales en parques y xardinos. Una mención estreme merez el casu de Narcissus (el narcisu), cultiváu en delles partes del mundu como ornamental en xardinos y tamién pa flor de corte.
Les afilantoideas entienden una sola especie, Aphyllanthes monspeliensis, distribuyida nes zones grebes del oeste del Mediterraneu. La planta ta entendida por agrupamientos d'inflorescencies escaposas, los escapos son en realidá'l principal órganu fotosintéticu, una y bones les fueyes escariosas na base nun son fotosintétiques. La inflorescencia tien poques flores, con tépalos usualmente azules de tamañu medianu. La planta ye paecida a Sisyrinchium.
Les brodiaeoideas (antes familia de les temidáceas) entienden una docena de xéneros nativos del oeste de Norteamérica. Son bien similares a les alioideas, yá que son yerbes perennes con una inflorescencia umbelada a la fin d'un escapo, con flores bastante pequeñes, y tépalos más o menos conados dacuando con una corona, y l'ovariu ye súpero, pero escarecen del "golor aliáceo" típicu d'estes postreres, y tienen un cormo fibrosu, non un bulbu, y amás hai usualmente 4 o más bráctees nes inflorescencies que nun son como les 3 bráctees de les alioideas, que son más envolventes.
Les scilloideas (antes familia de les hiacintáceas) entienden de 770 a 1.000 especies, distribuyíes predominantemente en climes mediterráneos, especialmente Sudáfrica y el Mediterraneu hasta Asia Central y Birmania (dalgunes en Suramérica). Tienen 6 tépalos y 6 estambres con ovariu súpero, por eso soler añerar en Liliaceae. Caracterizar por ser herbales perennes con bulbu, con fueyes bastante carnoses y mucilaginosas que se disponen nuna roseta basal. Tienen compuestos venenosos, polo que les sos especies nun son comestibles. Dellos xéneros son bien populares en xardinería, como Hyacinthus (el conocíu xacintu), importante como llanta cultivada y como flor cortada.
Les agavoideas tán distribuyíes más o menos por tol mundu y son especialmente diverses en Méxicu, onde Agave tien importantes usos económicos (por casu ye utilizáu pa ellaborar tequila y mescal). Munchos miembros d'esta subfamilia son arborescentes, como'l árbol de Josué (una especie de Yucca). Les fueyes son munches vegaes grandes, xeromórficas, fibroses o raramente ensundioses, en rosetas na base o'l final de les cañes. Les flores presenten 6 tépalos y 6 estambres, y l'ovariu ye súpero o ínfero.
Les lomandroideas (antes familia de les laxmaniáceas) son predominantemente australianas. Tienen los tépalos persistentes nel frutu. La familia puede ser estremada en 3 grupos con calteres distintivos: el grupu Lomandra, el grupu Cordyline y el grupu Laxmannia.
Les asparagoideas tán distribuyíes pel Vieyu Mundu (Asparagus) y Méxicu (Hemiphylacus). Los órganos fotosintéticos esplanaos de Asparagus fueron suxetu de muncho discutiniu, sicasí, la mayoría de los autores considerar tarmos esplanaos. Ye interesáu que cañes fotosintétiques similares evolucionaren independientemente en Nolinoideae (antes Ruscaceae), subfamilia cola que les asparagoideas tán emparentaes. Asparagus tien fueyes amenorgaes a escames non fotosintétiques que lleven na so axila filoclados verdes, solitarios a abigarrados, circulares a esplanaos en seición tresversal, y el frutu ye una baga.
Les nolinoideas (antes familia de les ruscáceas), como güei definíes (nun sentíu más ampliu que'l tradicional), tán llargamente distribuyíes en rexones templaes a tropicales. Son herbales a arborescentes, dacuando tienen crecedera secundaria anómalu, dacuando los tarmos son esplanaos y fotosintéticos (son filoclados). Les flores bisexuales, de simetría radial, son pequeñes. Los 6 tépalos suelen tar conados na base polo que les flores tienen forma de campana o similares. Tienen 6 estambres y un ovariu súpero. El frutu usualmente ye una baga de poques granes.
La monofilia de Asparagales ta sostenida por analís cladísticos basaos en morfoloxía, ADNr 18S, y munches secuencies d'ADN (Chase et al. 1995a,[20]b,[21] 2000,[22] 2006,[23] Conran 1989,[24] Davis et al. 2004,[25] Graham et al. 2006,[26] Hilu et al. 2003,[27] Pires et al. 2006,[28] Rudall et al. 1997,[29] Källersjö et al. 1998,[30] Fay et al. 2000,[31] Soltis et al. 2000,[32] Stevenson et al. 2000,[33] McPherson y Graham 2001),[34] magar delles reconstrucciones filoxenétiques como la de Hilu et al. (2003),[27] con datos moleculares, suxurieron que Asparagales pue ser parafiléticu, con Orchidaceae separáu del restu.
Con al respective de les rellaciones de Asparagales col restu de los órdenes de monocotiledónees, ye'l clado hermanu del clado Commelinidae.
Fixeron analises morfolóxicos Chase et al. (1995b)[21] y Stevenson y Loconte (1995),[35] y nos dos siquier dalgunes de les families de Asparagales con crecedera secundaria anómalu formaben clados, magar tamién cayíen dientro d'estos clados dalgunos miembros que güei tán allugaos en Liliales, Zingiberales y Pandanales. Tamién ver Rudall (2002)[36] pa un trabayu recién de filoxenia del orde con datos morfolóxicos.
Les rellaciones dientro del orde fueron investigaes por Dahlgren et al. (1985),[6] Rudall y Cutler (1995),[37] Chase et al. (1995a,[20]b,[21] 1996),[38] Stevenson y Loconte (1995),[35] Rudall et al. (1997),[29] Fay et al. (2000),[31] McPherson y Graham (2001),[34] Graham et al. (2006)[26] y Pires et al. (2006).[28]
El cladograma resumíu, según Judd et al. 2007 (modificáu de Soltis et al. 2005,[39] Orchidaceae basal como n'APWeb 2008):[4]
Asparagales
Hypoxidaceae (tradicionalmente allugáu en Amaryllidaceae)
Xanthorrhoeaceae sensu llato
Hemerocallidoideae (tradicionalmente allugáu en Liliaceae)
Xanthorrhoeoideae (tradicionalmente allugáu en Liliaceae)
Asphodeloideae (tradicionalmente allugáu en Liliaceae)
Amaryllidaceae sensu llato
Agapanthoideae (tradicionalmente allugáu en Liliaceae)
Allioideae (tradicionalmente allugáu en Liliaceae)
Lomandroideae (antes familia Laxmanniaceae, tradicionalmente allugáu en Liliaceae)
Asparagoideae (tradicionalmente allugáu en Liliaceae)
Nolinolidease (antes familia Ruscaceae, tradicionalmente allugáu en Liliaceae)
Brodiaeoideae (antes familia Themidaceae, tradicionalmente allugáu en Liliaceae)
Scilloideae (antes Hyacinthaceae, tradicionalmente allugáu en Liliaceae)
Agavoideae (tradicionalmente allugáu en Amaryllidaceae o Liliaceae)
Un cladograma completu ye apurríu pol APWeb.[4]
Asparagales
Xanthorrhoeaceae sensu llato
Amaryllidaceae sensu llato
Brodiaeoideae = Themidaceae
Scilloideae = Hyacinthaceae
Agavoideae (Anemarrhena, Agave y allegaos, Hesperocallis, Behnia, Herreria y allegaos, Anthericum y allegaos)
Lomandroideae = Laxmanniaceae
Nolinoideae = Ruscaceae
L'árbol, colos nomes de los clados actualizaos según el APG III (2009),[1] ta basáu principalmente nos analises de Chase et al. (2000)[22] y Fay et al. (2000).[31] Estos estudios difieren pocu nos detalles, magar l'analís de Fay et al. (2000)[31] tien más nodos con bon sofitu nel nucleu de los Asparagales (lo cual nun ye sorprendente, yá que utilizaron el métodu de pesaje socesivu o "successive weighting"). Tamién ye largamente congruente con esta filoxenia la presentada por McPherson y Graham (2001),[34] magar el so muestreo ye enforma menor.
De los analís esprende un asemblaje basal parafiléticu, llamáu los "Baxos Asparagales" (sensu Rudall et al. 1997),[29] qu'inclúi a families como Orchidaceae, Hypoxidaceae, Iridaceae, Asphodeloideae, y Hemerocallidoideae. En contraste, los llamaos "Altos Asparagales" son un clado puramente dichu que s'esprende del restu, ya inclúi a Allioideae, Amaryllidoideae, Scillloideae = Hyacinthaceae, Agavoideae, Asparagoideae, y Nolinoideae = Ruscaceae. Munchos de los "Baxos Asparagales" caracterizar pola microsporogénesis simultánea (les 4 microsporas dixebrar a un tiempu ente sigo dempués de los dos divisiones meióticas). El patrón de microsporogénesis de los "Baxos Asparagales" ye apomórfico dientro de les monocotiledónees. El patrón de microsporogénesis de los "Altos Asparagales" sicasí, ye como aquellos de les monocotiledónees más basales. Rudall (2001,[40] ver tamién 2002b,[41] 2003a)[42] incluyó al caracter del ovariu ínfero como una sinapomorfía del orde, notando que nos "Altos Asparagales" podría haber una reversión al ovariu súpero, reversión acomuñada cola presencia de nectarios septales infraloculares (como en Xanthorrhoea y Johnsonia, dos xéneros de Hemerocallidoideae). Sicasí, Stevens (2001 d'equí p'arriba, APWeb)[4] nota que los ovarios súperos tamién apaecen distribuyíos tou a lo llargo de los "Baxos Asparagales", polo qu'ónde tienen d'allugase nel árbol les evoluciones de les distintes morfoloxíes del ovariu nun ta claru: la posición del ovariu paez un calter muncho más flexible (equí como nes demás anxospermes) de lo que se creía. Los "Altos Asparagales", por ser un grupu monofiléticu con fuerte sofitu nos analises moleculares d'ADN, son llamaos "Nucleu de los Asparagales" ("Core Asparagales") pol APWeb.[4]
Orchidaceae ye la familia más grande de toles anxospermes. Dellos autores (por casu Dahlgren et al. 1985)[6] reconocieron tres families d'orquídees, pero los analises moleculares d'ADN demostraron qu'estes families son polifiléticas y tendríen d'axuntase nun Orchidaceae sensu llato, que ye como se trata a la familia anguaño. Con al respective de la so rellación coles demás families de Asparagales, dellos trabayos recién suxuren que Orchidaceae ye hermana del restu de los Asparagales (por casu con sofitu bootstrap del 76 % en Graham et al. 2006,[26] un resultáu más o menos similar en Givnish et al. 2006b,[43] un sofitu más fuerte -del 96 al 99 %-, con un bon tamañu de muestreo y 7 xenes de dos compartimientos, en Pires et al. 2006,[28] y tamién un bon sofitu en Chase et al. 2005,[44] onde'l nodo dixebrando a les orquídees del restu de Asparagales tenía 91 % de sofitu bootstrap). Otros trabayos allugar en forma distinta nel árbol filoxenéticu: dacuando foi allugada como hermana de Boryaceae (por casu en Chase et al. 1995a[20] y en McPherson y Graham 2001,[34] magar con un sofitu más bien baxu). Sicasí Li y Zhou (2007)[45] atopen a Orchidaceae enfiñida nun clado parafiléticu Boryaceae-Hypoxidaceae. Per otru llau Rudall (2003a),[42] con datos morfolóxicos, suxure una rellación morfolóxica cercana ente Hypoxidaceae y Orchidaceae en particular. Janssen y Bremer (2004),[46] magar nun alluguen a Orchidales como hermanos del restu del orde, allugar temporalmente cerca del empiezu de la diverxencia dientro d'él (anque la topoloxía del árbol qu'utilizaron difier considerablemente en detalle del utilizáu equí). En resume, la posición de Orchidaceae como hermanu del restu de Asparagales, con Boryaceae como parte del clado qu'inclúi a Blandfordiaceae, paez la meyor hipótesis, y esta topoloxía ye la presentada equí. Nótese qu'esto camuda la caracterización de Asparagales, munchos de los sos calteres agora siendo reasitiaos al nodo subbasal nel clado (comparar la llista de apomorfías del APWeb[4] en versiones 7 y anteriores a esa del sitiu web). Un datu interesante ye que les orquídees escarecen de la mayoría de les sinapomorfíes non moleculares del orde: les granes de Orchidaceae nun tienen fitomelaninas por cuenta de la so naturaleza (son como un polvillo, como la mayoría de los micoparásitos), y los sos nectarios raramente tán nos septos de los ovarios. Les orquídees tienen microsporogénesis simultánea y ovariu ínfero, calteres que son típicos de los "Baxos Asparagales" de Rudall et al. 1997,[29] pero nun ta claru si estos calteres tendríen de considerase sinapomorfías del orde. La única sinapomorfía de Orchidaceae aparentemente ye la presencia de protocormo (la estructura producida pola crecedera de los sos embriones indiferenciados primero que los raigaños y tarmos desenvuélvanse). Les orquídees presenten l'androcéu y el xinecéu fundíos, pero esta carauterística ta presente namái na subfamilia Apostasioideae y namái en graos variables. Les granes como polvillo esvalixaes por vientu tán presentes na mayoría de les subfamilies, pero en dellos miembros de Cypripedioideae y Vanilloideae hai granes crustosas (probablemente acomuñaes a la so forma de dispersión, que nesti casu ye por páxaros y mamíferos que son atraíos pol so frutu carnosu que llelda in situ lliberando compuestos fragantes, por casu la vainillina). La diversificación en Orchidaceae ye remarcable, pero tien de ser comparada cola de los demás Asparagales, que son menos ricos n'especies pero bien variables morfolóxicamente.
Les rellaciones de Boryaceae (qu'entiende namái dos xéneros, Borya y Alania) col restu de les families de Asparagales permanecieron pocu clares mientres enforma tiempu. Les boryáceas son micorrícicas, pero non como les orquídees, sinón que les sos micorrizas son del tipu estandarizado vesicular-arbuscular (VA). Primeramente llegóse a pensar que Boryaceae yera un miembru d'Anthericaceae (Dahlgren et al. 1985,[6] Takhtajan 1997),[47] una familia que demostró ser desaxeradamente polifilética (Chase et al. 1996).[38] L'allugamientu de Orchidaceae como hermana de too el restu de Asparagales da-y un sofitu más o menos importante al allugamientu de Boryaceae como hermanu del clado qu'inclúi a Blandfordiaceae. Rudall (2003a)[42] tamién suxurió una rellación morfolóxica cercana ente Boryaceae y Blandfordiaceae. Sicasí hai que tener en cuenta que magar hai bon sofitu nel árbol de Chase et al. (2006)[23] p'asitiar a Orchidaceae como hermanu de toles demás Asparagales, Boryaceae ta allugáu darréu percima del clado qu'inclúi a Blandfordiaceae, magar con bien so sofitu.
Hypoxidaceae ye una familia que tradicionalmente fuera allugada en Amaryllidoideae. Lanariaceae col so únicu xéneru Lanaria nel pasáu foi consideráu un miembru de Haemodoraceae (Hutchinson 1967)[48] o Tecophilaeaceae (Dahlgren et al. 1985).[6] Estos dos families xuntu con Asteliaceae, Blandfordiaceae y probablemente tamién Boryaceae formen un clado, y salvu pol allugamientu de Boryaceae, les otres 4 tienen bon sofitu como clado nos analises moleculares. Una sinapomorfía potencial pa estes families ye la estructura del óvulu: siquier Asteliaceae, Blandfordiaceae, Lanariaceae y Hypoxidaceae tienen una constricción chalazal y una cofia nucelar ("nucellar cap"). Soltis et al. (2005), dexando fuera del clado a Boryaceae, suxurieron qu'estes families se fusionararan nun Hypoxidaceae sensu llato.
Dientro d'esti clado, Asteliaceae + Hypoxidaceae + Lanariaceae formen de la mesma un clado con sofitu del 100 % de bootstrap. La morfoloxía tamién aprove dalgún sofitu pa estes rellaciones: Asteliaceae y Hypoxidaceae formen rosetas cubiertes con pelos multicelulares ramificaos (tamién Lanariaceae tien pelos multicelulares), y tienen canales radiculares llenos de mucílagu (esti postreru caracter tamién presente en Lanariaceae), pero Blandfordia (únicu xéneru de Blandfordiaceae) nun comparte estos calteres. Amás, esti clado tien estomes paracíticos, la llámina de la fueya con una vena media distintiva, quiciabes el xinecéu más o menos ínfero, y una micropila biestomal. Asteliaceae + Hypoxidaceae tienen flavonoles, endosperma de parés delgaes, cotiledón non fotosintéticu, y la lígula llarga.
El siguiente nodo en Asparagales (toles families menos Orchidaceae y el clado mentáu) demostróse que ye robustu (81 % de sofitu jacknife en Soltis et al. 2000,[32] 100 % de sofitu bootstrap en Chase et al. 2005,[44] ver tamién Fay et al. 2000[31] usando datos de cuatro gen plastídicos, y Chase et al. 2000[22]), pero nun hai calteres fuera de los moleculares que xunan a esti clado.
La posición d'Ixioliriaceae y Tecophilaeaceae inda ye pocu clara. Dellos analises moleculares xunir nun clado, por casu recibe sofitu de leve a moderáu en Chase et al. (2000),[22] Pires et al. (2006),[28] y Givnish et al. (2006),[43] y en Graham et al. (2006)[26] recibe un sofitu más fuerte pero'l muestreo ye más bien probe. Nel analís de 3 xenes de Fay et al. (2000)[31] el clado tien un brazu bien llargu. Estos dos families comparten los cormos, la inflorescencia foliosa, y munches vegaes, una inflorescencia cuasi capitada, les fueyes de disposición espiralada y base envainadora, les flores más bien grandes, el verticilu esternu de tépalos mucronado a aristado, el perianto formando un tubu curtiu, el androcéu ensertáu na boca, y un númberu cromosómico x=12 (Soltis et al. 2005 y APWeb). Tamién s'atopen otres topoloxíes: en Janssen y Bremer (2004)[46] Ixioliriaceae diverxe considerablemente antes (magar ye axacente a) Tecophilaeaceae; y en Davis et al. (2004),[25] con un muestreo probe, atópase dalgún sofitu pa hermanar a Ixioliriaceae con Iridaceae, tamién en Chase et al. (2006)[23] atópase sofitu (esta vegada fuerte) pa esti parentescu, onde'l clado ye hermanu de Doryanthaceae (pero con sofitu baxu).
Les rellaciones de Doryanthaceae coles families que la arrodien nos analises filoxenéticos inda son pocu clares tantu nos analises morfolóxicos como nos moleculares. Chase et al. (2006)[23] atopar nel allugamientu recién mentáu, que nun ye la presentada equí. Pal allugamientu presentáu equí hai sofitu moderáu en Fay et al. (2000),[31] y un sofitu de 92 % en Graham et al. (2006).[26] Rudall (2003)[42] atopa una rellación morfolóxica cercana ente Doryanthaceae y Iridaceae.
El nodo que contién a Iridaceae y a tol restu de los Asparagales (Xeronemataceae, etc.) recibe un sofitu bastante fuerte en Fay et al. (2000).[31] El grupu tien delles carauterístiques moleculares de mancomún: perdió los telómeros de tipu Arabidopsis, ente que son comunes los telómeros de tipu humanu (TTAGGG).
Iridaceae ye una de les families más grandes y meyor estudiaes de Asparagales (por casu ver Goldblatt 1990,[49] 1991,[50] Goldblatt et al. 1998,[51] 2002,[52] Reeves et al. 2001).[53] Iridaceae ye distintiva ente los Asparagales por cuenta de la estructura única de la so inflorescencia (un ripidio) y la so combinación d'ovariu ínfero con 3 estambres, tamién son comunes na familia les fueyes unifaciales, ente que les fueyes bifaciales son la norma n'otros Asparagales.
El nodo que sigue al posible desprendimientu de Iridaceae ta bien sosteníu nos analises (Fay et al. 2000,[31] Chase et al. 2005).[44] Si un ovariu ínfero ye una sinapomorfía de Asparagales (Chase et al. 1995b)[21] entós esti nodo marcaría la transición a un ovariu súpero (anque s'atopen ovarios ínferos en Amaryllidoideae y en Yucca). Tolos clados d'esti nodo tienen nectarios septales infraloculares, lo que Rudall (2000)[54] interpreta como ovarios secundariamente súperos. Esta interpretación demuestra la reversibilidad d'esti calter que munches vegaes foi enfatizado na sistemática de monocotiledónees (por casu en Cronquist 1981,[55] Dahlgren et al. 1985).[6] Ye nesti nodo nel que se pierde'l xen mitocondrial rpl2 (o quiciabes nel siguiente, acordies con la distribución en Adams et al. 2002).[56]
Xeronemataceae col so únicu xéneru Xeronema foi primeramente consideráu cercanamente emparentáu con Phormium (usualmente Phormiaceae, Dahlgren et al. 1985),[6] pero magar hai semeyances superficiales (como les fueyes unifaciales), nun son idénticos nos detalles. Chase et al. (2000)[22] apocayá describió a Xeronemataceae, y los sos calteres varien según patrones que son xenerales en Asparagales.
El siguiente nodo del árbol (Xanthorrhoeaceae sensu llato + los "Altos Asparagales") tien fuerte sofitu en Fay et al. (2000)[31] y en Chase et al. (2000c).[57]
Hemerocallidoideae ta cercana a Asphodelaceoideae y Xanthorrhoeoideae, y los 3 clados tienen antraquinones, estiloides, y grana con cotiledón non fotosintéticu. Ver Kite et al. (2000)[58] pa la distribución de antraquinonas, McPherson et al. (2004)[59] pa los taxones que nun tienen el intrón 3'-rps12. Dientro d'esti clado, asocede crecedera secundaria (anómalu) en Aloe (delles especies llegando a tamaños enormes), Phormium y Xanthorrhoea. Les 3 families son por dellos autores axuntaes nun Xanthorrhoeaceae sensu llato (como n'APG III y nel APWeb, 2001 d'equí p'arriba),[4] anque Stevens nel APWeb remarca que la microsporogénesis ye variable nesti grupu. En Fay et al. (2000)[31] atópase fuerte sofitu pa esti clado. Sicasí, les rellaciones de parentescu dientro d'él entá nun tán esclariaes. Nel analís de 3 xenes de Chase et al. (2000)[22] hai un sofitu moderáu pa la rellación (Xanthorrhoeoideae + Asphodeloideae), qu'ente los calteres morfolóxicos atopa la presencia de engrosamiento secundariu, l'androcéu non adnato a los tépalos, la presencia de hipostasa ("hypostase"), y les granes anguloses. Esta rellación ye la presentada equí, sicasí'l sofitu mengua cuando s'inclúin los taxones onde falten delles secuencies de xenes, ver tamién Fay et al. 2000).[31] Sicasí, Devey et al. (2006)[60] atopen dalgún sofitu p'axuntar a Xanthorrhoeoideae con Hemerocallidoideae (ver tamién Pires et al. 2006),[28] ente que Chase et al. (2006)[23] suxuren un clado formáu por Asphodeloideae + Hemerocallidoideae (pero'l muestreo ye cuestionáu nel APWeb).[4] Con al respective de la morfoloxía, tanto Hemerocallidoideae como Xanthorrhoeoideae tienen ovarios que probablemente son secundariamente súperos, y tienen nectarios septales infraloculares (Rudall 2002,[36] 2003).[42] Rudall (2003)[42] suxure una rellación morfolóxica cercana ente Hemerocallidoideae y Asphodeloideae, y otra ente Xanthorrhoeoideae y Iridaceae.
El valor de les antraquinonas como caracter taxonómicu ye apreciáu agora, yá que Asphodeloideae tener pero enantes nun fuera allugada como cercana a les otres dos families de Xanthorrhoeaceae sensu llato sinón a Anthericaceae (cola cual agora saber que ta lejanamente rellacionada). Foi por cuenta de la presencia de antraquinonas que Asphodeloideae foi allugada dixebrada de Anthericaceae por Dahlgren et al. (1985),[6] amás de que Asphodeloideae tien microsporogénesis simultánea ente que Anthericaceae tien microsporogénesis socesiva. Asphodeloideae tien munchos xéneros con cariotipos bimodales distintivos (Chase et al. 2000c),[57] y esti caracter foi usáu de base pa suxurir qu'estos xéneros fueren dixebraos nuna familia Aloaceae (Dahlgren et al. 1985).[6] Sicasí los estudios moleculares caltuvieron que Asphodeloideae quedar na so circunscripción amplia.
Xanthorrhoeoideae antes incluyía otros taxones arborescentes como Kingia (que nun tien crecedera secundaria y agora ye allugáu en Dasypogonaceae) y Cordyline (col cual ta namái lejanamente emparentáu, ye un miembru de Lomandroideae, antes Laxmanniaceae).
Hemerocallidaceae munches vegaes entendía namái al xéneru Hemerocallis (Dahlgren et al. 1985),[6] pero yá dende la circunscripción del APG (1998),[7] la familia incluyía un gran númberu de xéneros, como Dianella, Johnsonia, y Phormium. Toos estos xéneros tienen polen tricotomosulcado, una rareza ente les monocotiledónees, y los estudios moleculares tempranos (Chase et al. 1995a)[20] fueron críticos en determinar si esti caracter yera importante pa estos xéneros (Rudall et al. 1997).[29] Nel APG III ye la subfamilia Hemerocallidoideae de Xanthorrhoeaceae sensu llato.
El restu de les families, que podríen axuntase nos clados Amaryllidaceae sensu llato y Asparagaceae sensu llato, denominar en xunto "Nucleu de los Asparagales" pol APWeb,[4] caracterizar por tener microsporogénesis socesiva (son los "Altos Asparagoides" de Rudall et al. 1997),[29] y quiciabes tamién por tener saponines esteroideas. Nos analises moleculares d'ADN ye un grupu con fuerte sofitu (por casu en Chase et al. 1995a,[20] Fay et al. 2000,[31] Chase et al. 2000c,[57] Graham et al. 2005).[61] Sicasí, la inclusión d'Aphyllanthoideae (en Asparagaceae sensu llato), tiende a menguar el sofitu (Graham et al. 2006).[26] Amás, hai pocu sofitu morfolóxicu especialmente pa les agrupaciones dientro de la área d'Asparagaceae.
Amaryllidaceae sensu llato (Allioideae, Amaryllidoideae y Agapanthoideae) ta axuntáu poles sos secuencies de ADNcp (Fay et al. 2000,[31] pero ver McPherson et al. 2004[59] y Thomas et al. 2005) y les sos secuencies ADNnr ITS para Amaryllidoideae + Agapanthoideae (Meerow et al. 2000.[62]
Dientro del clado, [Agapanthoideae [Allioideae + Amaryllidoideae]] ye suxuríu por Meerow et al. (1999),[63] Fay et al. (2000,[31] con fuerte sofitu), Givnish et al. (2006)[43] y Pires et al. (2006),[28] pero Meerow et al. (2000a)[64] atoparon a Agapanthoideae como hermana de Amaryllidoideae, magar con baxu sofitu.
El clado tien dellos calteres morfolóxicos: les inflorescencies son escaposas, umbeloides (construcción cimosa), xeneralmente con dos (o más) bráctees como espatas escariosas fundíes zarrándoles. Les fueyes son dísticas, los pedicelos non articulaos, l'estilu llargu, l'estigma secu, l'endosperma nuclear o helobial, ensin hipocótilo, quiciabes se debiera incluyir a los tépalos llibres, les anteres conadas na base, y les célules del tapete uninucleadas. (Allioideae + Amaryllidoideae) ten como posibles sinapomorfías el fechu de que son geofitas, el tarmu que ye un bulbu de ramificación simpodial, tunicado, y los raigaños contráctiles.
Les 7 families que formen el clado Asparagaceae sensu llato tán bien sosteníes nel analís de Fay et al. (2000,[31] ensin incluyir Hesperocallis), pero nun hai calteres morfolóxicos bien evidentes qu'axunten al clado, anque ye bon candidatu'l endosperma: helobial, de parés grueses, hemicelulósico. Inclusive les families en sentíu estrictu d'esti clado son difíciles de reconocer, porque nun tener sinapomorfías obvies, y les sos flores tienen el típicu aspeutu de les liliáceas (nesi sentíu'l APWeb[4] suxure qu'axuntales nun Asparagaceae sensu llato podría ser útil, como de fechu el APG III nel 2009 facer). Pa detalles de les sos rellaciones, ver Jang y Pfosser (2002)[65] y Bogler et al. (2006).[66]
En Asparagaceae sensu llato, a diferencia de Amaryllidaceae sensu llato, les inflorescencies son recímanos (magar Brodiaeoideae = Themidaceae tien inflorescencies umbeloides pero nesti casu, tienen bráctees per debaxo de toles flores, polo que les flores interprétense como recímanos entestaos). En Asparagaceae sensu llato hai munchos xéneros arborescentes (como Yucca, Agave, Cordyline), y tamién dalgunos con raigaños con crecedera secundaria (como Herreria, Thysanotus), y munchos son geofitas. Amaryllidaceae sensu llato ye muncho menos diversa, cola mayoría geofitas, bulboses, magar dalgunes son rizomatosas (como Agapanthus, Tulbaghia, Clivia). Un caracter morfolóxicu qu'axunta a les asparagáceas sensu llato ye la presencia d'una inflorescencia racimosa.
Brodiaeoideae = Themidaceae, Scilloideae = Hyacinthaceae, Aphyllanthoideae y Agavoideae formen un clado (ver Fay et al. 2000[31] pa toes menos Aphyllanthoideae), magar el sosten morfolóxicu pa esti grupu ye pocu claru. Esti clado puede ser hermanu del clado (Nolinoideae = Ruscaceae + Asparagoideae + Lomandroideae = Laxmanniaceae).
Les rellaciones ente Aphyllanthoideae, Brodiaeoideae = Themidaceae y Scilloideae = Hyacinthaceae quiciabes tengan de ser representaes como una tricotomía. Fay et al. (2000),[31] Pires et al. (2001),[67] y Pires y Sytsma (2002)[68] alderiquen la incertidume con al respective de les rellaciones de Brodiaeoideae. (Brodiaeoideae + Scilloideae) paecen tener un moderáu sosten en Fay y Chase (1996)[69] y en Meerow et al. (2000),[64] pero'l sosten ye baxu nel analís de 2 xenes de Jang y Pfosser (2002,[65] ensin incluyir Aphyllanthes), en Chase et al. (2006),[23] y en Pires et al. (2006).[28] Los dos clados comparten les saponines esteroideas, les fueyes de disposición espiralada, los pedicelos bracteados, l'óvulu anátropo, el endosperma helobial o nuclear, y el cotiledón non fotosintéticu. Pa dellos calteres más ver Fay y Chase (1996).[69] Estructures asemeyaes a laticíferos pueden tar presentes nos dos clados.
Aphyllanthes tien una caña bien llarga nel analís de 3 xenes de Fay et al. (2000),[31] y la so posición filoxenética ye pocu clara, pero'l so remoción de dellos analises mengua drásticamente el sofitu (Chase et al. 2006).[23] Una rellación de Aphyllantes como hermanu de Scilloideae foi atopada por McPherson y Graham (2001),[34] pero Pires et al. (2006)[28] atopar como hermanu de Lomandroideae, pero con sofitu baxu.
Les subfamilies Lomandroideae + Asparagoideae + Nolinoideae comparten les saponines esteroideas, los pedicelos articulaos, el frutu que ye una cápsula, l'endosperma helobial, de parés grueses, con puntuaciones, hemicelulósico. Hai sofitu moderáu pa esti clado nel árbol de 4 xenes del cloroplastu de Fay et al. (2000).[31]
Finalmente, Nolinoideae y Asparagoideae pueden tar rellacionaes, como s'evidencia polos sos frutos indehiscentes, les sos flores bastante pequeñes y el so númberu cromosómico x=10, esta rellación recibe prelilminar sofitu por dellos analises de secuencies d'ADN (Chase et al. 1995a,[20]b,[21] 1996,[38] 2000,[22] Fay et al. 2000[31]). Los frutos carnosos con granes ensin fitomelanines son comunes nesti clado, pero nun se conoz si esti caracter constitúi una apomorfía. Tienen frutos capsulares Hemiphylacus (hermanu del restu de Asparagoideae) y el clado (Comosperma + Eriospermum), que ye hermanu del restu de Nolinoideae, polo que puede ser que'l frutu carnosu seya deriváu dientro del clado.
Pal desarrollu de los filoclados en Asparagus, Ruscus, y allegaos, ver Cooney-Sovetts y Sattler (1987).[70]
El sistema APG de 1998[7] y el APG II del 2003[8] reconocieron a Hesperocallidaceae col so únicu xéneru Hesperocallis, pero foi allugáu en Agavaceae polos postreros analises moleculares d'ADN (Pires et al. 2004),[71] por eso la familia foi incluyida en Agavaceae en APG III del 2009[1] y nel APWeb.[4]
L'orde foi reconocíu pol APG III (2009),[1] el Linear APG III (2009)[2] asignó-y los númberos de familia 62 a 75. L'orde yá fuera reconocíu pol APG II (2003).[8]
L'orde, como anguaño ta circunscripto, consiste en 14 families y aproximao 26.800 especies. Les llendes de delles families inda son pocu escamplaes, y como s'hai descripto na seición de filoxenia, dellos botánicos propunxeron que se combinen munches de les families qu'equí se reconocen. Por casu, el APG III y el APWeb traten a Alliaceae, Amaryllidaceae y Agapanthaceae, que xuntos formen un clado, como una única familia: Amaryllidaceae sensu llato. Tamién combinen Xanthorrhoeaceae, Asphodelaceae y Hemerocallidaceae nun Xanthorrhoeaceae sensu llato, y a Agavaceae, Asparagoideae, Hyacinthaceae, Ruscaceae, Themidaceae, y unes poques families pequeñes sían combinaes, cola familia resultante llamada Asparagaceae sensu llato. Esta decisión ye taxonómicamente arbitraria, magar factores como la facilidá de la reconocencia y el nivel de sosten de la monofilia del grupu tendríen de ser consideraos.
Les families o subfamilies más representaes son Orchidaceae, Hypoxidaceae, Iridaceae, Amaryllidoideae, Allioideae, Scilloideae, Lomandroideae, Agavoideae, Asparagoideae, Convallariaceae, Asphodeloideae, y Hemerocallidoideae.
La llista de families según APG III (2009),[1] xuntu col númberu de familia según el Linear APG III (2009),[2] y dalgunes de les families tradicionalmente reconocíes y güei abandonaes, col so eventual destín, se listan de siguío:
Sinónimos (APWeb):[4] Asparagineae J. Presl, Asphodelineae Thorne & Reveal, Hyacinthineae Link, Iridineae Engler - Agavales Hutchinson, Alliales Traub, Amaryllidales Bromhead, Asphodelales Doweld, Asteliales Dumortier, Hypoxidales Reveal & Doweld, Iridales Rafinesque, Ixiales Lindley, Narcissales Dumortier, Orchidales Rafinesque, Tecophilaeales Reveal, Xanthorrhoeales Reveal & Doweld - Iridanae Doweld, Orchidanae Doweld - Orchididae Heintze - Crinopsida Horaninov, Orchidopsida Bartling
Nel Angiosperm Phylogeny Website (xunetu de 2008)[4] les families son:
El sistema APG II del 2003,[8] dispon al Orde Asparagales nel clado de les Monocotiledónees y utiliza la siguiente circunscripción:
Nota: "+..." = Familia que opcionalmente puede segregase de la familia precedente. Esto ye que, so esti sistema un taxónomo puede, por casu, allugar al xéneru Hemerocallis na familia Hemerocallidaceae o bien, na familia Xanthorrhoeaceae sensu llato (en sentíu ampliu).
Hesperocallidaceae col so únicu xéneru Hesperocallis foi allugáu en Agavaceae polos postreros analises moleculares d'ADN, por eso foi incluyida en Agavaceae n'APWeb.[4]
El Sistema APG de 1998[7] tamién dispunxo al orde Asparagales dientro del clado de les monocotiledónees, pero utilizó una circunscripción distinta de les families.
Nel volume 1998 (a,[14]b[72]) del Sistema Kubitzki utilizóse la siguiente circunscripción de families dientro de Asparagales:
Nel Sistema Dahlgren (1985)[6] l'orde Asparagales foi dispuestu nel Superorde Lilianae dientro de la Subclase Liliidae (= Monocotiledónees) de la Clase Magnoliopsida (= Anxospermes), y utilizó la siguiente circunscripción:
El sistema de Cronquist (1981)[55] nun reconoz al orde Asparagales y dispon a munchos de los sos taxones dientro del orde Liliales (na subclase Liliidae, dientro de la clase Liliopsida). Dellos xéneros taben inclusive dispuestos dientro de la familia Liliaceae.
El Sistema Wettstein, que data de 1935, tampoco reconocía al orde Asparagales, y disponía a munches de les sos especies dientro del orde Liliflorales na clase de les monocotiledónees.
El grupu troncal Asparagales data d'unos 122 millones d'años hasta'l presente, el grupu corona Asparagales d'unos 119 millones d'años hasta'l presente (Janssen y Bremer 2004),[46] magar Wikström et al. (2001)[73] suxurieron edaes de 107-98 y 101-94 millones d'años hasta'l presente respeutivamente. Hai que tener en cuenta que nel últimu estudiu utilizóse una topoloxía de Asparagales que difier sustancialmente de l'aceptada equí (especialmente cerca de la base del árbol).
La diverxencia del clado que contién a toles families salvu Hypoxidaceae sensu llato, Boryaceae y Orchidaceae, foi datáu nunos 84 millones d'años hasta'l presente (Eguiarte 1995).[74]
El "Nucleu de los Asparagales" dixebrar de Xanthorrhoeaceae sensu llato fai unos 93 millones d'años, la diverxencia dientro del clado empieza fai unos 91 millones d'años (Janssen y Bremer 2004),[46] sicasí en Wikström et al. (2001)[73] envalórense feches de 61-54 y 58-51 millones d'años hasta'l presente respeutivamente.
El grupu troncal Alliaceae sensu llato ta datáu nunos 91 millones d'años hasta'l presente, la diverxencia dientro del grupu corona Alliaceae sensu llato empieza fai unos 87 millones d'años (Janssen y Bremer 2004).[46] Nótese que los fungos qu'infecten a Allium y otres aliáceas son abondo distintes de los que s'atopen en Amaryllidaceae (ver por casu Savile 1962).[75]
El grupu troncal Asparagaceae sensu llato ta datáu nunos 91 millones d'años, la diverxencia dientro del grupu corona Asparagaceae sensu llato empieza fai unos 89 millones d'años (Janssen y Bremer 2004).[46] Eguiarte (1995),[74] sicasí, suxure que Agavaceae-Nolinaceae diverxeron namái fai unos 47 millones d'años.
Disposición de les fueyes basales (Agapanthus).
Disposición de les fueyes na estremidá del tueru en Yucca.
Vezu de crecedera de Rhodohypoxis.
Flor de Cattleya (Orchidaceae).
Flor de Hymenocallis (Amaryllidaceae).
Flores de Convallaria.
Asparagales ye'l nome d'un taxón de plantes perteneciente a la categoría taxonómica d'orde, utilizáu en sistemes de clasificación modernos como'l sistema de clasificación APG III del 2009 y l'APWeb, y ta circunscripto obligadamente siquier pola familia Asparagaceae. Asparagales ye un orde namái apocayá reconocíu nos sistemes de clasificación, y munches de les sos families pertenecíen al antiguu orde Liliales. Liliales fuera un orde bien ampliu onde práuticamente s'allugaben toles monocotiledónees de tépalos vistosos y ensin almidón nel endosperma, que fuera malo d'estremar en families porque los sos calteres morfolóxicos nun se daben en patrones que delimitaran grupos claramente. Col tiempu empezar a reconocer l'orde Asparagales, especialmente cuando los analises moleculares d'ADN indicaron que Liliales tenía de ser estremáu siquier en Liliales, Asparagales y Dioscoreales. Asparagales ye un orde abondo grande, consiste en 14 families y unes 26.000 especies. L'orde ta claramente circunscripto sobre la base de los analises de secuencies d'ADN pero ye dificultosu de definir morfolóxicamente, yá que ye unu de los grupos de monocotiledónees estructuralmente más diversu. Asina, l'orde inclúi yerbes de non más de 15 cm hasta delles especies de Agave que superen los 10 m d'altor. Nesti sentíu, la so separación del orde Liliales ye particularmente problemática.
Una de les carauterístiques que definen a esti orde, y posible apomorfía (amás de los datos moleculares), ye la presencia de fitomelanina, un pigmentu escuru presente nes cubiertes seminales. Les fitomelaninas atópase na mayoría de les families de Asparagales, perdiéndose aparentemente en dellos taxones, siendo Orchidaceae l'exemplu más conspicuo nesi aspeutu.
La mayoría de les especies de Asparagales son plantes yerbácees perennes, magar hai dellos exemplos de plantes trepadores (ciertes especies de Asparagus, subfamilia Asparagoideae) y maderices (dellos Aloe, subfamilia Asphodeloideae). Cuasi toles especies d'esti orde producen un recímanu apertáu de fueyes na base de les plantes (son "plantes en roseta"), otres lleven esa mesma disposición de fueyes nel estremu d'un tueru (Yucca), en dellos pocos casos les fueyes producir a lo llargo del tarmu. Les flores frecuentemente tópense na estremidá del tarmu.
Dientro de les monocotiledónees y dende el puntu de vista económicu, l'orde Asparagales ye'l segundu n'importancia dempués del orde Poales (el cual inclúi a los camperes y los ceberaes). L'orde contién munches especies d'importancia hortícola (cebolla, ayu, espárragu, puerru), en floricultura (fresia, gladiolu, iris, munchos miembros d'orquídees), xardinería (Hemerocallis, Convallaria, Agapanthus) y especies (vainilla).
Dende'l puntu de vista de la so taxonomía, Asparagales ye un orde suxuríu por Huber (1977), depués adoptáu por Dahlgren et al. (1985), y reafianzado en 1998 pol Grupu de Filoxenia de les Anxospermes (n'inglés orixinal, "Angiosperm Phylogeny Group", sigles APG), retomáu nel 2003 pol APG II, y nel 2009 pol APG III), y tamién pol APWeb (2001 hasta agora). Les delimitaciones de les families de Asparagales sufrieron una serie de cambeos n'años recién, y entá se precisen más estudios por que se estabilice la so circunscripción y les sos rellaciones.
Quşqonmazçiçəklilər (lat. Asparagales) — birləpəlilərə aid bitki sırası.
Bu sıranın tanınmış nümayındələri soğan və qulançar cinslərinin nümayəndələridir.
Qulançarkimilər sırasına aid inciçiçəyi, nərgizgülü, amarillis, səhləb kimi çoxlu dekorativ bitki cinsləri var.
Aqava, əzvay, yukka kimi otaq bitkiləri də bu sıraya aiddir.
Les asparagals (Asparagales) són un ordre de les plantes amb flor (Magnoliophyta). Aquest ordre inclou la família de les Asparagaceae i d'altres famílies que no sempre n'han format part, que segons el sistema de classificació presenten una marcada variació en el seu emplaçament.
El sistema de classificació APG II del 2003, inclou l'ordre Asparagales dintre del clade monocots i presenta aquestes agrupacions:
Nota: "+ ..." = família separada opcional, pot separar-se de la família precedent.
El sistema APG II ha agrupat algunes de les famílies del primer sistema APG al mateix temps que reconeix una alternativa permetent que petites famílies siguin segregades. Seguint aquest nou sistema de classificació el taxonomista pot, per exemple, escollir entre incloure els lliris de Sant Joan (Hemerocallis) a la família Hemerocallidaceae o a la família Xanthorrhoeaceae sense que la seva decisió deixi de ser correcta.
El primer sistema APG del 1998 també incloïa l'ordre Asparagales dintre del clade monocots però presentava aquestes agrupacions:
El sistema Kubitzki de classificació de les plantes vasculars utilitzava aquestes agrupacions:
El sistema Dahlgren incloïa l'ordre Asparagales dins del superordre Lilianae a la subclasse Liliidae [= monocotiledònies] de la classe Magnoliopsida [= angiospermes] i presentava aquestes agrupacions:
El sistema Cronquist de classificació no reconeixia l'ordre de les Asparagales i la majoria de les plantes eren dins l'ordre Liliales (a la subclasse Liliidae de la classe Liliopsida [= monocotiledònies]). Alguns gèneres eren inclosos dins la família Liliaceae.
El sistema Wettstein, a l'última revisió de 1935, no reconeixia aquest ordre, i classificava moltes de les plantes en l'ordre Liliiflorae de la classe de les Monocotiledònies.
Un possible arbre filogenètic dels asparagals, incloent-hi aquelles famílies recentment reduïdes a subfamílies, es mostra a sota.[1]
AsparagalesHemerocallidoideae (= Hemerocallidaceae)
Xanthorrhoeoideae (= Xanthorrhoeaceae s.s.)
Asphodeloideae (= Asphodelaceae)
Agapanthoideae (= Agapanthaceae)
Allioideae (= Alliaceae s.s.)
Amaryllidoideae (= Amaryllidaceae s.s.)
Aphyllanthoideae (= Aphyllanthaceae)
Brodiaeoideae (= Themidaceae)
Scilloideae (= Hyacinthaceae)
Agavoideae (= Agavaceae)
Lomandroideae (= Laxmanniaceae)
Asparagoideae (= Asparagaceae s.s.)
Nolinoideae (= Ruscaceae)
Les asparagals (Asparagales) són un ordre de les plantes amb flor (Magnoliophyta). Aquest ordre inclou la família de les Asparagaceae i d'altres famílies que no sempre n'han format part, que segons el sistema de classificació presenten una marcada variació en el seu emplaçament.
El sistema de classificació APG II del 2003, inclou l'ordre Asparagales dintre del clade monocots i presenta aquestes agrupacions:
ordre Asparagales família Alliaceae [+ família Agapanthaceae] [+ família Amaryllidaceae] família Asparagaceae [+ família Agavaceae] [+ família Aphyllanthaceae] [+ família Hesperocallidaceae] [+ família Hyacinthaceae] [+ família Laxmanniaceae] [+ família Ruscaceae] [+ família Themidaceae] família Asteliaceae família Blandfordiaceae família Boryaceae família Doryanthaceae família Hypoxidaceae família Iridaceae família Ixioliriaceae família Lanariaceae família Orchidaceae família Tecophilaeaceae família Xanthorrhoeaceae [+ família Asphodelaceae] [+ família Hemerocallidaceae] família XeronemataceaeNota: "+ ..." = família separada opcional, pot separar-se de la família precedent.
El sistema APG II ha agrupat algunes de les famílies del primer sistema APG al mateix temps que reconeix una alternativa permetent que petites famílies siguin segregades. Seguint aquest nou sistema de classificació el taxonomista pot, per exemple, escollir entre incloure els lliris de Sant Joan (Hemerocallis) a la família Hemerocallidaceae o a la família Xanthorrhoeaceae sense que la seva decisió deixi de ser correcta.
El primer sistema APG del 1998 també incloïa l'ordre Asparagales dintre del clade monocots però presentava aquestes agrupacions:
ordre Asparagales família Agapanthaceae família Agavaceae família Alliaceae família Amaryllidaceae família Anemarrhenaceae família Anthericaceae família Aphyllanthaceae família Asparagaceae família Asphodelaceae família Asteliaceae família Behniaceae família Blandfordiaceae família Boryaceae família Convallariaceae família Doryanthaceae família Hemerocallidaceae família Herreriaceae família Hyacinthaceae família Hypoxidaceae família Iridaceae família Ixioliriaceae família Lanariaceae família Laxmanniaceae família Orchidaceae família Tecophilaeaceae família Themidaceae família Xanthorrhoeaceae família XeronemataceaeEl sistema Kubitzki de classificació de les plantes vasculars utilitzava aquestes agrupacions:
order Asparagales família Agapanthaceae família Agavaceae família Alliaceae família Amaryllidaceae família Anemarrhenaceae família Anthericaceae família Aphyllanthaceae família Asparagaceae família Asphodelaceae família Asteliaceae família Behniaceae família Blandfordiaceae família Boryaceae família Convallariaceae família Doryanthaceae família Dracaenaceae família Eriospermaceae família Hemerocallidaceae família Herreriaceae família Hyacinthaceae família Hostaceae família Hypoxidaceae família Iridaceae família Ixioliriaceae família Johnsoniaceae família Lanariaceae família Lomandraceae família Nolinaceae família Orchidaceae família Ruscaceae família Tecophilaeaceae família Themidaceae família XanthorrhoeaceaeEl sistema Dahlgren incloïa l'ordre Asparagales dins del superordre Lilianae a la subclasse Liliidae [= monocotiledònies] de la classe Magnoliopsida [= angiospermes] i presentava aquestes agrupacions:
order Asparagales família Agavaceae família Alliaceae família Amaryllidaceae família Anthericaceae família Aphyllanthaceae família Asparagaceae família Asphodelaceae família Asteliaceae família Blandfordiaceae família Calectasiaceae família Convallariaceae família Cyanastraceae família Dasypogonaceae família Doryanthaceae família Dracaenaceae família Eriospermaceae família Hemerocallidaceae família Herreriaceae família Hostaceae família Hyacinthaceae família Hypoxidaceae família Ixioliriaceae família Lanariaceae família Luzuriagaceae família Nolinaceae família Philesiaceae família Phormiaceae família Ruscaceae família Tecophilaeaceae família XanthorrhoeaceaeEl sistema Cronquist de classificació no reconeixia l'ordre de les Asparagales i la majoria de les plantes eren dins l'ordre Liliales (a la subclasse Liliidae de la classe Liliopsida [= monocotiledònies]). Alguns gèneres eren inclosos dins la família Liliaceae.
El sistema Wettstein, a l'última revisió de 1935, no reconeixia aquest ordre, i classificava moltes de les plantes en l'ordre Liliiflorae de la classe de les Monocotiledònies.
Chřestotvaré (Asparagales), zvané také chřestokvěté, jsou velkým řádem jednoděložných rostlin. Zástupci tohoto řádu se vyvinuli ve střední křídě.
Pojetí řádu se v průběhu vývoje systematiky značně měnilo. Starší taxonomické systémy řád neznaly a velkou část zástupců řadily do široce pojaté čeledi liliovité (Liliaceae). Některé novější systémy, jako např. Tachtadžjanův systém mají řád mnohem úžeji pojatý a rozlišují ještě další jako Orchidales, Amaryllidales atd., jiné jako třeba Cronquistův systém ho neznají vůbec. Podle systému APG II sem patří zhruba 14-25 čeledí, podle toho, jak jsou čeledi pojaté; asi 1122 rodů a 26000 druhů (velkou většinu z toho však zabírají vstavačovité).
vstavačovité (Orchidaceae)
boriovité (Boryaceae)
blandfordiovité (Blandfordiaceae)
bičulovité (Asteliaceae)
tvrzeňovité (Hypoxidaceae)
ixiolirionovité (Ixioliriaceae)
šafránovcovité (Tecophilaeaceae)
hůlovcovité (Doryanthaceae)
kosatcovité (Iridaceae)
asfodelovité (Asphodelaceae)
amarylkovité (Amaryllidaceae)
chřestovité (Asparagaceae)
Jsou to převážně vytrvalé byliny, často s cibulemi, hlízami či oddenky. Ale někdy mohou být i stromkovitého charakteru s atypickým druhotným tloustnutím, viz třeba žlutokap. Značně jsou mezi nimi zastoupeny sukulenty, jako agáve, aloe aj., zato vodních rostlin je zde málo. Listy jsou jednoduché, často v přízemních nebo vrcholových růžicích, čepele jsou převážně čárkovité až kopinaté, méně často jiného tvaru, žilnatina je převážně souběžná. Někdy mohou být listy funkčně nahrazeny fylokladii, viz třeba chřest nebo listnatec. Květy se nejčastěji skládají ze 6 okvětních lístků, tyčinek bývá nejčastěji 6, gyneceum se skládá ze 3 plodolistů, semeník může být svrchní i spodní. Plodem je nejčastěji tobolka, méně bobule aj.
Agapanthus africanus
Yucca brevifolia
Anthericum ramosum
Narcissus pseudonarcissus
Chřestotvaré (Asparagales), zvané také chřestokvěté, jsou velkým řádem jednoděložných rostlin. Zástupci tohoto řádu se vyvinuli ve střední křídě.
Asparges-ordenen (Asparagales) er en planteorden med talrige familier.
Nedenfor nævnes nogle af de mere kendte familier, som er repræsenteret ved arter, der er vildtvoksende eller dyrkede i Danmark.
Familier
Die Spargelartigen (Asparagales) sind eine Ordnung der Monokotyledonen mit 14 teils umfangreichen Familien. Eine große Anzahl von Arten werden als Zierpflanzen für Parks, Gärten, Räume und als Schnittblumen genutzt. Viele Arten werden als Heilpflanzen verwendet. Wenige Arten sind wichtige Nahrungspflanzen, beispielsweise Spargel sowie einige Vertreter der Gattung Lauch, wie Zwiebel, Lauch oder Knoblauch.
Es sind meistens ausdauernde krautige Pflanzen oder Sukkulenten. Wenn überhaupt, dann ist nur anomales sekundäres Dickenwachstum (beispielsweise viele Arten der Grasbäume (Xanthorrhoea)) vorhanden, deshalb nennt man verholzende Arten hier baumförmig und nicht Baum. Die krautigen Taxa haben oft Speicherorgane (Zwiebeln oder Rhizome). Bei den Asparagales kommen Sukkulenten vor, bei den Liliales nie.
Die Blütenstände sind meist traubig. Die Blüten sind dreizählig. Drei Fruchtblätter sind zu einem meist oberständigen, seltener unterständigen Fruchtknoten verwachsen. Die Nektarsekretion erfolgt meist durch Septalnektarien am Fruchtknoten. Die Früchte sind in Fächer gegliederte Kapseln oder Beeren. Die Samenoberflächen sind meist durch Phytomelane schwarz.
Oft enthalten die Zellen Kristallsand in Form von Raphiden, das sind nadelfeine Kristalle (aus Calciumoxalat).
Molekulargenetische Untersuchungen haben in den letzten gut zehn Jahren dazu geführt, dass die Familiengrenzen sich stark verschoben haben. Einige Familien, beispielsweise die Grasbaumgewächse, sind stark erweitert worden, einige – darunter auch traditionelle Familien wie die Tagliliengewächse oder Lauchgewächse – wurden aufgelöst. Eine ausführliche Begründung dieser neuen Klassifikation sowie die Gegenüberstellung von traditionellen und neuen Familiennamen geben Chase u. a. 2009.[1] Der Hintergrund war vor allem, eine pragmatische und in der Lehre leicht zu vermittelnde Familien-Klassifikation zu schaffen.
Zur Ordnung der Spargelartigen (Asparagales) gehören folgende 14 Familien[2]
mit folgendem Verwandtschaftsverhältnissen:[3][4]
Asparagales„höhere“ Asparagales
Die Spargelartigen (Asparagales) sind eine Ordnung der Monokotyledonen mit 14 teils umfangreichen Familien. Eine große Anzahl von Arten werden als Zierpflanzen für Parks, Gärten, Räume und als Schnittblumen genutzt. Viele Arten werden als Heilpflanzen verwendet. Wenige Arten sind wichtige Nahrungspflanzen, beispielsweise Spargel sowie einige Vertreter der Gattung Lauch, wie Zwiebel, Lauch oder Knoblauch.
Asparagales is the name o an order o plants, uised in modren classification seestems such as the APG III seestem (which is uised throughoot this airticle).[5] The order taks its name frae the faimily Asparagaceae an is placed in the monocots. The order haes anly recently been recognized in classification seestems. It wis first put forward bi Huber in 1977[6] and later taken up in the Dahlgren seestem o 1985.[7] Afore this, mony o its faimilies wur assigned tae the auld order Liliales: a very lairge order containin almaist aw monocots wi colourful tepals a wioot starch in thair endosperm. DNA sequence analysis indicatit that Liliales should be dividit intae at least Liliales, Asparagales an Dioscoreales. The boondaries o the Asparagales an o its faimilies hae unnergone a series o chynges in recent years; future research mey lead tae further chynges an ultimately greater stability.
Data relatit tae Asparagales at Wikispecies
De Asparagales zyn in de biologische taxonomie een orde van de bedektzoadign of bloeinde plantn. De noame is gevormd uut de familienoame Asparagaceae.
Indêlienge van de orde volgens 't APG III-systeim (2009):
D'Asparagales sinn zesumme mat de Liliales an der Flora vu Mëtteleuropa déi wichtegst Uerdnung bannent der Ënnerklass vun de Liliidae.
Zur Uerdnung vun den Asparagales gehéiere 16 bis 24 Famillje mat 1.122 Gattungen, an iwwer 26.000 Aarten:
Asparagales è n'òrdini di chianti chî ciuri.
Lu Sistema APG II dû 2003, cunzigghia sta classificazzioni:
Lu Sistema Cronquist nun ricanuscìu st'òrdini.
L' Asparagales hè un ordine di a classa di i Liliopsida chi cumprende isse famiglie:
Asparagales is the name o an order o plants, uised in modren classification seestems such as the APG III seestem (which is uised throughoot this airticle). The order taks its name frae the faimily Asparagaceae an is placed in the monocots. The order haes anly recently been recognized in classification seestems. It wis first put forward bi Huber in 1977 and later taken up in the Dahlgren seestem o 1985. Afore this, mony o its faimilies wur assigned tae the auld order Liliales: a very lairge order containin almaist aw monocots wi colourful tepals a wioot starch in thair endosperm. DNA sequence analysis indicatit that Liliales should be dividit intae at least Liliales, Asparagales an Dioscoreales. The boondaries o the Asparagales an o its faimilies hae unnergone a series o chynges in recent years; future research mey lead tae further chynges an ultimately greater stability.
D'Asparagales sinn zesumme mat de Liliales an der Flora vu Mëtteleuropa déi wichtegst Uerdnung bannent der Ënnerklass vun de Liliidae.
Koma çavmarkan (Asparagales) komeke riwekan e, li ser yekqalikan tê hejmartin. 14 famîleyên riwekan di vê komê de cih digirin. Pirraniya riwekên wê bo baxçevaniyê tên çandin. Hin jî tên xwarin. Bo mînak çavmark, sîr, pirasa, pîvaz.
Koma çavmarkan (Asparagales) komeke riwekan e, li ser yekqalikan tê hejmartin. 14 famîleyên riwekan di vê komê de cih digirin. Pirraniya riwekên wê bo baxçevaniyê tên çandin. Hin jî tên xwarin. Bo mînak çavmark, sîr, pirasa, pîvaz.
A spargeloortagen (Asparagales) san en kategorii faan plaanten mä 14 familin. Enkelt slacher, so üs spargel of pore wurd tu iidjen brükt.
A spargeloortagen (Asparagales) san en kategorii faan plaanten mä 14 familin. Enkelt slacher, so üs spargel of pore wurd tu iidjen brükt.
Спаржакветныя (Asparagales) — парадак кветкавых (Angiosperms) расьлінаў, які зьмяшчае 14 сямействаў і каля 35900 відаў[1].
Спаржакветныя (Asparagales) — парадак кветкавых (Angiosperms) расьлінаў, які зьмяшчае 14 сямействаў і каля 35900 відаў.
Шпаргловидни (науч. Asparagales) — ред во поткласата Liliidae и вклучува тревести, монокотиледонски цветни растенија кои се нашироко распространети. Сите претставници на овој ред поседуваат паралелна нерватура на листовите. Цветовите се исклучително разновидни по својата форма и боја. Името на редот е дадено по родот Asparagus (шпаргла).
Редот Шпаргловидни ги вклучува следните фамилии:
Asparagales (asparagoid lilies) is an order of plants in modern classification systems such as the Angiosperm Phylogeny Group (APG) and the Angiosperm Phylogeny Web. The order takes its name from the type family Asparagaceae and is placed in the monocots amongst the lilioid monocots. The order has only recently been recognized in classification systems. It was first put forward by Huber in 1977 and later taken up in the Dahlgren system of 1985 and then the APG in 1998, 2003 and 2009. Before this, many of its families were assigned to the old order Liliales, a very large order containing almost all monocots with colorful tepals and lacking starch in their endosperm. DNA sequence analysis indicated that many of the taxa previously included in Liliales should actually be redistributed over three orders, Liliales, Asparagales, and Dioscoreales. The boundaries of the Asparagales and of its families have undergone a series of changes in recent years; future research may lead to further changes and ultimately greater stability. In the APG circumscription, Asparagales is the largest order of monocots with 14 families, 1,122 genera, and about 36,000 species.
The order is clearly circumscribed on the basis of molecular phylogenetics, but it is difficult to define morphologically since its members are structurally diverse. Most species of Asparagales are herbaceous perennials, although some are climbers and some are tree-like. The order also contains many geophytes (bulbs, corms, and various kinds of tuber). According to telomere sequence, at least two evolutionary switch-points happened within the order. The basal sequence is formed by TTTAGGG like in the majority of higher plants. Basal motif was changed to vertebrate-like TTAGGG and finally, the most divergent motif CTCGGTTATGGG appears in Allium. One of the defining characteristics (synapomorphies) of the order is the presence of phytomelanin, a black pigment present in the seed coat, creating a dark crust. Phytomelanin is found in most families of the Asparagales (although not in Orchidaceae, thought to be the sister-group of the rest of the order).
The leaves of almost all species form a tight rosette, either at the base of the plant or at the end of the stem, but occasionally along the stem. The flowers are not particularly distinctive, being 'lily type', with six tepals and up to six stamina.
The order is thought to have first diverged from other related monocots some 120–130 million years ago (early in the Cretaceous period), although given the difficulty in classifying the families involved, estimates are likely to be uncertain.
From an economic point of view, the order Asparagales is second in importance within the monocots only to the order Poales (which includes grasses and cereals). Species are used as food and flavourings (e.g. onion, garlic, leek, asparagus, vanilla, saffron), in medicinal or cosmetic applications (Aloe), as cut flowers (e.g. freesia, gladiolus, iris, orchids), and as garden ornamentals (e.g. day lilies, lily of the valley, Agapanthus).


Although most species in the order are herbaceous, some no more than 15 cm high, there are a number of climbers (e.g., some species of Asparagus), as well as several genera forming trees (e.g. Agave, Cordyline, Yucca, Dracaena, Aloe ), which can exceed 10 m in height. Succulent genera occur in several families (e.g. Aloe).
Almost all species have a tight cluster of leaves (a rosette), either at the base of the plant or at the end of a more-or-less woody stem as with Yucca. In some cases, the leaves are produced along the stem. The flowers are in the main not particularly distinctive, being of a general 'lily type', with six tepals, either free or fused from the base and up to six stamina. They are frequently clustered at the end of the plant stem.
The Asparagales are generally distinguished from the Liliales by the lack of markings on the tepals, the presence of septal nectaries in the ovaries, rather than the bases of the tepals or stamen filaments, and the presence of secondary growth. They are generally geophytes, but with linear leaves, and a lack of fine reticular venation.
The seeds characteristically have the external epidermis either obliterated (in most species bearing fleshy fruit), or if present, have a layer of black carbonaceous phytomelanin in species with dry fruits (nuts). The inner part of the seed coat is generally collapsed, in contrast to Liliales whose seeds have a well developed outer epidermis, lack phytomelanin, and usually display a cellular inner layer.
The orders which have been separated from the old Liliales are difficult to characterize. No single morphological character appears to be diagnostic of the order Asparagales.
As circumscribed within the Angiosperm Phylogeny Group system Asparagales is the largest order within the monocotyledons, with 14 families, 1,122 genera and about 25,000–42,000 species, thus accounting for about 50% of all monocots and 10–15% of the flowering plants (angiosperms).[9][10] The attribution of botanical authority for the name Asparagales belongs to Johann Heinrich Friedrich Link (1767–1851) who coined the word 'Asparaginae' in 1829 for a higher order taxon that included Asparagus[11] although Adanson and Jussieau had also done so earlier (see History). Earlier circumscriptions of Asparagales attributed the name to Bromhead (1838), who had been the first to use the term 'Asparagales'.[4]
The type genus, Asparagus, from which the name of the order is derived, was described by Carl Linnaeus in 1753, with ten species.[12] He placed Asparagus within the Hexandria Monogynia (six stamens, one carpel) in his sexual classification in the Species Plantarum.[13] The majority of taxa now considered to constitute Asparagales have historically been placed within the very large and diverse family, Liliaceae. The family Liliaceae was first described by Michel Adanson in 1763,[14] and in his taxonomic scheme he created eight sections within it, including the Asparagi with Asparagus and three other genera.[15] The system of organising genera into families is generally credited to Antoine Laurent de Jussieu who formally described both the Liliaceae and the type family of Asparagales, the Asparagaceae, as Lilia and Asparagi, respectively, in 1789.[16] Jussieu established the hierarchical system of taxonomy (phylogeny), placing Asparagus and related genera within a division of Monocotyledons, a class (III) of Stamina Perigynia[17] and 'order' Asparagi, divided into three subfamilies.[18] The use of the term Ordo (order) at that time was closer to what we now understand as Family, rather than Order.[19][20] In creating his scheme he used a modified form of Linnaeus' sexual classification but using the respective topography of stamens to carpels rather than just their numbers. While De Jussieu's Stamina Perigynia also included a number of 'orders' that would eventually form families within the Asparagales such as the Asphodeli (Asphodelaceae), Narcissi (Amaryllidaceae) and Irides (Iridaceae), the remainder are now allocated to other orders. Jussieu's Asparagi soon came to be referred to as Asparagacées in the French literature (Latin: Asparagaceae).[21] Meanwhile, the 'Narcissi' had been renamed as the 'Amaryllidées' (Amaryllideae) in 1805, by Jean Henri Jaume Saint-Hilaire, using Amaryllis as the type species rather than Narcissus, and thus has the authority attribution for Amaryllidaceae.[22] In 1810, Brown proposed that a subgroup of Liliaceae be distinguished on the basis of the position of the ovaries and be referred to as Amaryllideae[23] and in 1813 de Candolle described Liliacées Juss. and Amaryllidées Brown as two quite separate families.[24]
The literature on the organisation of genera into families and higher ranks became available in the English language with Samuel Frederick Gray's A natural arrangement of British plants (1821).[25] Gray used a combination of Linnaeus' sexual classification and Jussieu's natural classification to group together a number of families having in common six equal stamens, a single style and a perianth that was simple and petaloid, but did not use formal names for these higher ranks. Within the grouping he separated families by the characteristics of their fruit and seed. He treated groups of genera with these characteristics as separate families, such as Amaryllideae, Liliaceae, Asphodeleae and Asparageae.[26]

The circumscription of Asparagales has been a source of difficulty for many botanists from the time of John Lindley (1846), the other important British taxonomist of the early nineteenth century. In his first taxonomic work, An Introduction to the Natural System of Botany (1830)[27] he partly followed Jussieu by describing a subclass he called Endogenae, or Monocotyledonous Plants (preserving de Candolle's Endogenæ phanerogamæ)[28] divided into two tribes, the Petaloidea and Glumaceae. He divided the former, often referred to as petaloid monocots, into 32 orders, including the Liliaceae (defined narrowly), but also most of the families considered to make up the Asparagales today, including the Amaryllideae.
By 1846, in his final scheme[29] Lindley had greatly expanded and refined the treatment of the monocots, introducing both an intermediate ranking (Alliances) and tribes within orders (i.e. families). Lindley placed the Liliaceae within the Liliales, but saw it as a paraphyletic ("catch-all") family, being all Liliales not included in the other orders, but hoped that the future would reveal some characteristic that would group them better. The order Liliales was very large and had become a used to include almost all monocotyledons with colourful tepals and without starch in their endosperm (the lilioid monocots). The Liliales was difficult to divide into families because morphological characters were not present in patterns that clearly demarcated groups. This kept the Liliaceae separate from the Amaryllidaceae (Narcissales). Of these Liliaceae[30] was divided into eleven tribes (with 133 genera) and Amaryllidaceae[31] into four tribes (with 68 genera), yet both contained many genera that would eventually segregate to each other's contemporary orders (Liliales and Asparagales respectively). The Liliaceae would be reduced to a small 'core' represented by the tribe Tulipae, while large groups such Scilleae and Asparagae would become part of Asparagales either as part of the Amaryllidaceae or as separate families. While of the Amaryllidaceae, the Agaveae would be part of Asparagaceae but the Alstroemeriae would become a family within the Liliales.
The number of known genera (and species) continued to grow and by the time of the next major British classification, that of the Bentham & Hooker system in 1883 (published in Latin) several of Lindley's other families had been absorbed into the Liliaceae.[32] They used the term 'series' to indicate suprafamilial rank, with seven series of monocotyledons (including Glumaceae), but did not use Lindley's terms for these. However they did place the Liliaceous and Amaryllidaceous genera into separate series. The Liliaceae[33] were placed in series Coronariae, while the Amaryllideae[34] were placed in series Epigynae. The Liliaceae now consisted of twenty tribes (including Tulipeae, Scilleae and Asparageae), and the Amaryllideae of five (including Agaveae and Alstroemerieae). An important addition to the treatment of the Liliaceae was the recognition of the Allieae[35] as a distinct tribe that would eventually find its way to the Asparagales as the subfamily Allioideae of the Amaryllidaceae.
The appearance of Charles Darwin's Origin of Species in 1859 changed the way that taxonomists considered plant classification, incorporating evolutionary information into their schemata. The Darwinian approach led to the concept of phylogeny (tree-like structure) in assembling classification systems, starting with Eichler.[36] Eichler, having established a hierarchical system in which the flowering plants (angiosperms) were divided into monocotyledons and dicotyledons, further divided into former into seven orders. Within the Liliiflorae were seven families, including Liliaceae and Amaryllidaceae. Liliaceae included Allium and Ornithogalum (modern Allioideae) and Asparagus.[37]
Engler, in his system developed Eichler's ideas into a much more elaborate scheme which he treated in a number of works including Die Natürlichen Pflanzenfamilien (Engler and Prantl 1888)[38] and Syllabus der Pflanzenfamilien (1892–1924).[39] In his treatment of Liliiflorae the Liliineae were a suborder which included both families Liliaceae and Amaryllidaceae. The Liliaceae[40] had eight subfamilies and the Amaryllidaceae[41] four. In this rearrangement of Liliaceae, with fewer subdivisions, the core Liliales were represented as subfamily Lilioideae (with Tulipae and Scilleae as tribes), the Asparagae were represented as Asparagoideae and the Allioideae was preserved, representing the alliaceous genera. Allieae, Agapantheae and Gilliesieae were the three tribes within this subfamily.[42] In the Amaryllidacea, there was little change from the Bentham & Hooker. A similar approach was adopted by Wettstein.[43]

In the twentieth century the Wettstein system (1901–1935) placed many of the taxa in an order called 'Liliiflorae'.[44] Next Johannes Paulus Lotsy (1911) proposed dividing the Liliiflorae into a number of smaller families including Asparagaceae.[45] Then Herbert Huber (1969, 1977), following Lotsy's example, proposed that the Liliiflorae be split into four groups including the 'Asparagoid' Liliiflorae.[46][47]
The widely used Cronquist system (1968–1988)[48][49][50] used the very broadly defined order Liliales.
These various proposals to separate small groups of genera into more homogeneous families made little impact till that of Dahlgren (1985) incorporating new information including synapomorphy. Dahlgren developed Huber's ideas further and popularised them, with a major deconstruction of existing families into smaller units. They created a new order, calling it Asparagales. This was one of five orders within the superorder Liliiflorae.[51] Where Cronquist saw one family, Dahlgren saw forty distributed over three orders (predominantly Liliales and Asparagales).[52][53] Over the 1980s, in the context of a more general review of the classification of angiosperms, the Liliaceae were subjected to more intense scrutiny. By the end of that decade, the Royal Botanic Gardens at Kew, the British Museum of Natural History and the Edinburgh Botanical Gardens formed a committee to examine the possibility of separating the family at least for the organization of their herbaria. That committee finally recommended that 24 new families be created in the place of the original broad Liliaceae, largely by elevating subfamilies to the rank of separate families.[54]
The order Asparagales as currently circumscribed has only recently been recognized in classification systems, through the advent of phylogenetics. The 1990s saw considerable progress in plant phylogeny and phylogenetic theory, enabling a phylogenetic tree to be constructed for all of the flowering plants. The establishment of major new clades necessitated a departure from the older but widely used classifications such as Cronquist and Thorne based largely on morphology rather than genetic data. This complicated discussion about plant evolution and necessitated a major restructuring.[55] rbcL gene sequencing and cladistic analysis of monocots had redefined the Liliales in 1995.[56][57] from four morphological orders sensu Dahlgren. The largest clade representing the Liliaceae, all previously included in Liliales, but including both the Calochortaceae and Liliaceae sensu Tamura. This redefined family, that became referred to as core Liliales, but corresponded to the emerging circumscription of the Angiosperm Phylogeny Group (1998).[58]
The 2009 revision of the Angiosperm Phylogeny Group system, APG III, places the order in the clade monocots.[59]
From the Dahlgren system of 1985 onwards, studies based mainly on morphology had identified the Asparagales as a distinct group, but had also included groups now located in Liliales, Pandanales and Zingiberales.[60] Research in the 21st century has supported the monophyly of Asparagales, based on morphology, 18S rDNA, and other DNA sequences,[61][62][63][64][65] although some phylogenetic reconstructions based on molecular data have suggested that Asparagales may be paraphyletic, with Orchidaceae separated from the rest.[66] Within the monocots, Asparagales is the sister group of the commelinid clade.[55]
This cladogram shows the placement of Asparagales within the orders of Lilianae sensu Chase & Reveal (monocots) based on molecular phylogenetic evidence.[67][59][68][69] The lilioid monocot orders are bracketed, namely Petrosaviales, Dioscoreales, Pandanales, Liliales and Asparagales.[70] These constitute a paraphyletic assemblage, that is groups with a common ancestor that do not include all direct descendants (in this case commelinids as the sister group to Asparagales); to form a clade, all the groups joined by thick lines would need to be included. While Acorales and Alismatales have been collectively referred to as "alismatid monocots" (basal or early branching monocots), the remaining clades (lilioid and commelinid monocots) have been referred to as the "core monocots".[71] The relationship between the orders (with the exception of the two sister orders) is pectinate, that is diverging in succession from the line that leads to the commelinids.[68] Numbers indicate crown group (most recent common ancestor of the sampled species of the clade of interest) divergence times in mya (million years ago).[69]
Lilianae sensu Chase & Reveal (monocots) 131[67] 122 120Dioscoreales 115
Pandanales 91
Liliales 121
121Asparagales 120
commelinids 118A phylogenetic tree for the Asparagales, generally to family level, but including groups which were recently and widely treated as families but which are now reduced to subfamily rank, is shown below.[9][1]
Asparagales Hypoxidaceae s.l. AsphodelaceaeHemerocallidoideae (= Hemerocallidaceae)
Xanthorrhoeoideae (= Xanthorrhoeaceae s.s.)
Asphodeloideae (= Asphodelaceae)
'core' Asparagales Amaryllidaceae s.l.Agapanthoideae (= Agapanthaceae)
Allioideae (= Alliaceae s.s.)
Amaryllidoideae (= Amaryllidaceae s.s.)
Asparagaceae s.l.Aphyllanthoideae (= Aphyllanthaceae)
Brodiaeoideae (= Themidaceae)
Scilloideae (= Hyacinthaceae)
Agavoideae (= Agavaceae)
Lomandroideae (= Laxmanniaceae)
Asparagoideae (= Asparagaceae s.s.)
Nolinoideae (= Ruscaceae)
The tree shown above can be divided into a basal paraphyletic group, the 'lower Asparagales (asparagoids)', from Orchidaceae to Asphodelaceae,[72] and a well-supported monophyletic group of 'core Asparagales' (higher asparagoids), comprising the two largest families, Amaryllidaceae sensu lato and Asparagaceae sensu lato.[1]
Two differences between these two groups (although with exceptions) are: the mode of microsporogenesis and the position of the ovary. The 'lower Asparagales' typically have simultaneous microsporogenesis (i.e. cell walls develop only after both meiotic divisions), which appears to be an apomorphy within the monocots, whereas the 'core Asparagales' have reverted to successive microsporogenesis (i.e. cell walls develop after each division).[61] The 'lower Asparagales' typically have an inferior ovary, whereas the 'core Asparagales' have reverted to a superior ovary. A 2002 morphological study by Rudall treated possessing an inferior ovary as a synapomorphy of the Asparagales, stating that reversions to a superior ovary in the 'core Asparagales' could be associated with the presence of nectaries below the ovaries.[73] However, Stevens notes that superior ovaries are distributed among the 'lower Asparagales' in such a way that it is not clear where to place the evolution of different ovary morphologies. The position of the ovary seems a much more flexible character (here and in other angiosperms) than previously thought.[1]
The APG III system when it was published in 2009, greatly expanded the families Xanthorrhoeaceae, Amaryllidaceae, and Asparagaceae.[9] Thirteen of the families of the earlier APG II system were thereby reduced to subfamilies within these three families. The expanded Xanthorrhoeaceae is now called "Asphodelaceae". The APG II families (left) and their equivalent APG III subfamilies (right) are as follows:
Orchidaceae is possibly the largest family of all angiosperms (only Asteraceae might - or might not - be more speciose) and hence by far the largest in the order. The Dahlgren system recognized three families of orchids, but DNA sequence analysis later showed that these families are polyphyletic and so should be combined. Several studies suggest (with high bootstrap support) that Orchidaceae is the sister of the rest of the Asparagales.[63][64][65][74] Other studies have placed the orchids differently in the phylogenetic tree, generally among the Boryaceae-Hypoxidaceae clade.[75][61][56][76][77] The position of Orchidaceae shown above seems the best current hypothesis,[1] but cannot be taken as confirmed.
Orchids have simultaneous microsporogenesis and inferior ovaries, two characters that are typical of the 'lower Asparagales'. However, their nectaries are rarely in the septa of the ovaries, and most orchids have dust-like seeds, atypical of the rest of the order. (Some members of Vanilloideae and Cypripedioideae have crustose seeds, probably associated with dispersal by birds and mammals that are attracted by fermenting fleshy fruit releasing fragrant compounds, e.g. vanilla.)
In terms of the number of species, Orchidaceae diversification is remarkable. However, although the other Asparagales may be less rich in species, they are more variable morphologically, including tree-like forms.
The four families excluding Boryaceae form a well-supported clade in studies based on DNA sequence analysis. All four contain relatively few species, and it has been suggested that they be combined into one family under the name Hypoxidaceae sensu lato.[78] The relationship between Boryaceae (which includes only two genera, Borya and Alania), and other Asparagales has remained unclear for a long time. The Boryaceae are mycorrhizal, but not in the same way as orchids. Morphological studies have suggested a close relationship between Boryaceae and Blandfordiaceae.[61] There is relatively low support for the position of Boryaceae in the tree shown above.[63]
The relationship shown between Ixioliriaceae and Tecophilaeaceae is still unclear. Some studies have supported a clade of these two families,[63] others have not.[75] The position of Doryanthaceae has also varied, with support for the position shown above,[64] but also support for other positions.[63]
The clade from Iridaceae upwards appears to have stronger support. All have some genetic characteristics in common, having lost Arabidopsis-type telomeres.[79] Iridaceae is distinctive among the Asparagales in the unique structure of the inflorescence (a rhipidium), the combination of an inferior ovary and three stamens, and the common occurrence of unifacial leaves whereas bifacial leaves are the norm in other Asparagales.
Members of the clade from Iridaceae upwards have infra-locular septal nectaries, which Rudall interpreted as a driver towards secondarily superior ovaries.[73]
The next node in the tree (Xanthorrhoeaceae sensu lato + the 'core Asparagales') has strong support.[80] 'Anomalous' secondary thickening occurs among this clade, e.g. in Xanthorrhoea (family Asphodelaceae) and Dracaena (family Asparagaceae sensu lato), with species reaching tree-like proportions.
The 'core Asparagales', comprising Amaryllidaceae sensu lato and Asparagaceae sensu lato, are a strongly supported clade,[64] as are clades for each of the families. Relationships within these broadly defined families appear less clear, particularly within the Asparagaceae sensu lato. Stevens notes that most of its subfamilies are difficult to recognize, and that significantly different divisions have been used in the past, so that the use of a broadly defined family to refer to the entire clade is justified.[1] Thus the relationships among subfamilies shown above, based on APWeb as of December 2010, is somewhat uncertain.
Several studies have attempted to date the evolution of the Asparagales, based on phylogenetic evidence. Earlier studies[81][82] generally give younger dates than more recent studies,[75][83] which have been preferred in the table below.
A 2009 study suggests that the Asparagales have the highest diversification rate in the monocots, about the same as the order Poales, although in both orders the rate is little over half that of the eudicot order Lamiales, the clade with the highest rate.[83]
The taxonomic diversity of the monocotyledons is described in detail by Kubitzki.[84][85] Up-to-date information on the Asparagales can be found on the Angiosperm Phylogeny Website.[1]
The APG III system's family circumscriptions are being used as the basis of the Kew-hosted World Checklist of Selected Plant Families.[86] With this circumscription, the order consists of 14 families (Dahlgren had 31)[51] with approximately 1120 genera and 26000 species.[1]
Order Asparagales Link
The earlier 2003 version, APG II, allowed 'bracketed' families, i.e. families which could either be segregated from more comprehensive families or could be included in them. These are the families given under "including" in the list above. APG III does not allow bracketed families, requiring the use of the more comprehensive family; otherwise the circumscription of the Asparagales is unchanged. A separate paper accompanying the publication of the 2009 APG III system provided subfamilies to accommodate the families which were discontinued.[87] The first APG system of 1998 contained some extra families, included in square brackets in the list above.
Two older systems which use the order Asparagales are the Dahlgren system[60] and the Kubitzki system.[84] The families included in the circumscriptions of the order in these two systems are shown in the first and second columns of the table below. The equivalent family in the modern APG III system (see below) is shown in the third column. Note that although these systems may use the same name for a family, the genera which it includes may be different, so the equivalence between systems is only approximate in some cases.
The Asparagales include many important crop plants and ornamental plants. Crops include Allium, Asparagus and Vanilla, while ornamentals include irises, hyacinths and orchids.[10]
Asparagales (asparagoid lilies) is an order of plants in modern classification systems such as the Angiosperm Phylogeny Group (APG) and the Angiosperm Phylogeny Web. The order takes its name from the type family Asparagaceae and is placed in the monocots amongst the lilioid monocots. The order has only recently been recognized in classification systems. It was first put forward by Huber in 1977 and later taken up in the Dahlgren system of 1985 and then the APG in 1998, 2003 and 2009. Before this, many of its families were assigned to the old order Liliales, a very large order containing almost all monocots with colorful tepals and lacking starch in their endosperm. DNA sequence analysis indicated that many of the taxa previously included in Liliales should actually be redistributed over three orders, Liliales, Asparagales, and Dioscoreales. The boundaries of the Asparagales and of its families have undergone a series of changes in recent years; future research may lead to further changes and ultimately greater stability. In the APG circumscription, Asparagales is the largest order of monocots with 14 families, 1,122 genera, and about 36,000 species.
The order is clearly circumscribed on the basis of molecular phylogenetics, but it is difficult to define morphologically since its members are structurally diverse. Most species of Asparagales are herbaceous perennials, although some are climbers and some are tree-like. The order also contains many geophytes (bulbs, corms, and various kinds of tuber). According to telomere sequence, at least two evolutionary switch-points happened within the order. The basal sequence is formed by TTTAGGG like in the majority of higher plants. Basal motif was changed to vertebrate-like TTAGGG and finally, the most divergent motif CTCGGTTATGGG appears in Allium. One of the defining characteristics (synapomorphies) of the order is the presence of phytomelanin, a black pigment present in the seed coat, creating a dark crust. Phytomelanin is found in most families of the Asparagales (although not in Orchidaceae, thought to be the sister-group of the rest of the order).
The leaves of almost all species form a tight rosette, either at the base of the plant or at the end of the stem, but occasionally along the stem. The flowers are not particularly distinctive, being 'lily type', with six tepals and up to six stamina.
The order is thought to have first diverged from other related monocots some 120–130 million years ago (early in the Cretaceous period), although given the difficulty in classifying the families involved, estimates are likely to be uncertain.
From an economic point of view, the order Asparagales is second in importance within the monocots only to the order Poales (which includes grasses and cereals). Species are used as food and flavourings (e.g. onion, garlic, leek, asparagus, vanilla, saffron), in medicinal or cosmetic applications (Aloe), as cut flowers (e.g. freesia, gladiolus, iris, orchids), and as garden ornamentals (e.g. day lilies, lily of the valley, Agapanthus).
Asparagaloj (latine: Asparagales) estas ordo de plantoj el la klaso de Unukotiledonaj plantoj. La nomon la ordo havas de la genro kaj specio asparago. Entute la ordo konsistas el 16 ĝis 24 familioj kun 1122 genroj kaj pli ol 26.000 specioj.
Inter alie cepo kaj ajlo, sed ankaŭ aloo kaj draceno, hiacinto, narciso kaj neĝborulo estas anoj de la planta parencaro.
Asparagaloj (latine: Asparagales) estas ordo de plantoj el la klaso de Unukotiledonaj plantoj. La nomon la ordo havas de la genro kaj specio asparago. Entute la ordo konsistas el 16 ĝis 24 familioj kun 1122 genroj kaj pli ol 26.000 specioj.
Inter alie cepo kaj ajlo, sed ankaŭ aloo kaj draceno, hiacinto, narciso kaj neĝborulo estas anoj de la planta parencaro.
Asparagales es un taxón de plantas perteneciente a la categoría taxonómica de orden, utilizado en sistemas de clasificación modernos como el sistema de clasificación APG III del 2009[3] y el APWeb,[4] y está circunscripto obligadamente al menos por la familia Asparagaceae. Asparagales es un orden solo recientemente reconocido en los sistemas de clasificación, y muchas de sus familias pertenecían al antiguo orden Liliales. Liliales había sido un orden muy amplio donde prácticamente se ubicaban todas las monocotiledóneas de tépalos vistosos y sin almidón en el endosperma, que había sido difícil de dividir en familias porque sus caracteres morfológicos no se daban en patrones que delimitaran grupos claramente. Con el tiempo se empezó a reconocer el orden Asparagales, especialmente cuando los análisis moleculares de ADN indicaron que Liliales debía ser dividido al menos en Liliales, Asparagales y Dioscoreales. Asparagales es un orden bastante grande, consiste en 14 familias y unas 26 000 especies. El orden está claramente circunscripto sobre la base de los análisis de secuencias de ADN pero es dificultoso de definir morfológicamente, ya que es uno de los grupos de monocotiledóneas estructuralmente más diverso. Así, el orden incluye hierbas de no más de 15 cm hasta algunas especies de Agave que superan los 10 metros de altura. En este sentido, su separación del orden Liliales es particularmente problemática.
Una de las características que definen a este orden, y posible apomorfía (además de los datos moleculares), es la presencia de fitomelanina, un pigmento oscuro presente en las cubiertas seminales. Las fitomelaninas se encuentran en la mayoría de las familias de Asparagales, habiéndose perdido aparentemente en algunos taxones, siendo Orchidaceae el ejemplo más conspicuo en ese aspecto.
La mayoría de las especies de Asparagales son plantas herbáceas perennes, si bien hay algunos ejemplos de plantas trepadoras (ciertas especies de Asparagus, subfamilia Asparagoideae) y leñosas (algunos Aloe, subfamilia Asphodeloideae). Casi todas las especies de este orden producen un racimo apretado de hojas en la base de las plantas (son "plantas en roseta"), otras llevan esa misma disposición de hojas en el extremo de un tronco (Yucca), en algunos pocos casos las hojas se producen a lo largo del tallo. Las flores frecuentemente se hallan en la extremidad del tallo.
Dentro de las monocotiledóneas y desde el punto de vista económico, el orden Asparagales es el segundo en importancia después del orden Poales (el cual incluye a los pastos y los cereales). El orden contiene muchas especies de importancia hortícola (cebolla, ajo, espárrago, puerro), en floricultura (freesia, gladiolo, iris, muchos miembros de orquídeas), jardinería (Hemerocallis, Convallaria, Agapanthus) y especias (vainilla).
Desde el punto de vista de su taxonomía, Asparagales es un orden sugerido por Huber (1977),[5] luego adoptado por Dahlgren et al. (1985),[6] y reafianzado en 1998 por el Grupo de Filogenia de las Angiospermas (en inglés original, "Angiosperm Phylogeny Group", siglas APG),[7] retomado en el 2003 por el APG II,[8] y en el 2009 por el APG III),[1] y también por el APWeb (2001 hasta ahora).[4] Las delimitaciones de las familias de Asparagales han sufrido una serie de cambios en años recientes, y aún se necesitan más estudios para que se estabilice su circunscripción y sus relaciones.
Grupo de las agaves, ajos, aloes, cebollas, dracaenas, espárragos, gladiolos, hemerocallis, iris, narcisos, orquídeas, puerros, vainilla,... Es una planta herbácea, rizomatosa, perennifolia, de aspecto muy variable y con varias subespecies. Tiene 6-20 hojas acintadas y arqueadas de hasta 70 x 5 cm, las cuales son tóxicas. Florece desde finales de otoño a principios de primavera, produciendo un tallo floral erecto, cilíndrico, de hasta un metro de altura, que termina en una inflorescencia más o menos esférica de hasta unas 30 flores. Estas flores miden hasta 5 cm de diámetro, tienen forma de trompeta y 6 tépalos azulados, en ocasiones blancos.
Es una especie utilizada con frecuencia en jardinería.
Procede originalmente del Natal y el Cabo de Buena Esperanza, en Sudáfrica. Actualmente se ha asilvestrado en gran parte del planeta.
Se confunde muy a menudo con su pariente Agapanthus africanus, una planta muy escasa en cultivo y muy difícil de cultivar. La práctica totalidad de los ejemplares que veamos en el comercio etiquetados como Agapanthus africanus son en realidad Agapanthus praecox. Estas dos especies son las únicas perennifolias del género, las restantes son caducifolias.
Es un Agave de gran tamaño, que forma grandes rosetas de hasta 3 x 5 metros, con hojas carnosas de hasta 2 x 25 cm, estrechadas hacia la base pero sin formar un peciolo, más anchas en la zona media y terminando en una punta cilíndrica endurecida, afilada, de hasta 4 cm, pinchosa. Las hojas frecuentemente se doblan cerca de la mitad de su longitud, de forma que el ápice queda dirigido hacia el suelo. El margen de la hoja, especialmente en la zona media, tiene grandes aguijones más o menos curvados, marrones. Las rosetas de hojas acumulan energía durante años para producir una descomunal inflorescencia que puede superar los 8 metros de altura, sostenida por un tronco cilíndrico, con brácteas esparcidas más o menos escuamiformes. La inflorescencia se divide en ramas laterales no ramificadas salvo en su ápice, donde forman una especie de agregado de flores con forma de paraguas, con las flores todas casi en un mismo plano, dirigidas hacia arriba. Cada flor mide 30-40 mm y tiene tépalos de 17-26{esd}}mm, estrechamente triangular-oblongos, erectos, de ápice obtuso, amarillos con la zona apical parda. El fruto es una cápsula trígona.
El carácter inusual de un crecimiento secundario anómalo con un meristema en hileras superpuestas ("etágeno") está prácticamente confinado a los géneros de este clado (fuera de este clado, está presente solo en Dioscorea). Algunas de estas especies, de los géneros Agave, Aloe, Cordyline, Dracaena, Nolina, y Yucca, se vuelven árboles cortos y macizos, generalmente (pero no siempre) con ramificación limitada (por ejemplo Aloe, Dracaena, Yucca). Otros géneros como Aphyllanthes y Lomandra, tienen este mismo tipo de crecimiento secundario pero confinado a sus tallos subterráneos.
Otros caracteres apomórficos del orden son: la presencia de ácido chelidónico, quizás la de velamen, quizás la de las células hipodérmicas de las raíces dimórficas, las anteras más largas que anchas, las células del tapete bi- a tetranucelares, la microsporogénesis simultánea (pero ver en apartado de Filogenia), la presencia de estilo, las semillas con exotesta, el tegmen no persistente, el endosperma helobial, y la pérdida del gen mitocondrial sdh3.
Si la microsporogénesis simultánea es una apomorfía del orden, en el "Núcleo de los Asparagales" aparentemente cambió a sucesiva. Hay taxones de clados más basales donde la microsporogénesis es sucesiva, donde los detalles de la formación de la pared son similares a los de aquellos miembros del núcleo de los Asparagales que fueron estudiados: son láminas centrífugas ("centrifugal cell plates"). Pero en aquellos miembros basales donde la microsporogénesis es simultánea, la formación de las láminas también puede ser centrífuga (Nadot et al. 2006).[9]
Algunos autores consideran al ovario ínfero una sinapomorfía del orden, pero si lo fuera, fue perdido tantas veces que resulta ser un carácter muy lábil (dentro del orden como en el resto de las angiospermas), por lo que la posición del ovario dentro del orden no tiene valor predictivo.
Tépalos de 3 trazas se encuentran en Orchidaceae, Amaryllidoideae, Iridaceae, Asphodeloideae (pero no en Kniphofia, Asphodelus), Agavoideae, Agapanthoideae, y Hemerocallidoideae; se encuentran tépalos de 1 traza en Nolinoideae (pero no en Maianthemum stellatum), Allioideae, Aphyllanthoideae, y Asparagoideae. Scilloideae tiene tépalos con los dos tipos de vasculatura, incluso se encuentran tépalos de 5 trazas en los verticilos externos de Urginea.
Pareciera que Asparagales representa una situación dinámica en la cual se mantiene una variedad de motivos teloméricos en baja cantidad de copias, en lugar de la repetición dominante de tipo Arabidopsis. Parece que en algunos casos, uno de los tipos que estaba en baja cantidad de copias es amplificado y reemplaza al tipo estándar. Por ejemplo, se sabía desde hace muchos años que Allium no poseía las repeticiones teloméricas estandarizadas de tipo Arabidopsis, y luego fue descubierto (Adams et al. 2001)[10] que Aloe tampoco las tenía, por lo que se predijo que los géneros intermedios tampoco las tendrían: en un análisis de representantes de todos los Asparagales se descubrió que, salvo Ornithogalum, ninguno tenía las repeticiones de Arabidopsis. Más adelante (Sykorova et al. 2003)[11] se descubrió que Ornithogalum y varios otros géneros habían reemplazado las repeticiones estándar de las plantas por unas de tipo humano, y que prácticamente todos los géneros de Asparagales tienen al menos algunas copias de las repeticiones teloméricas de tipo humano, como también una variedad de otras repeticiones ubicadas en las terminaciones de sus cromosomas. La presencia y la composición de esta variedad de repeticiones teloméricas poseen un gran valor para discernir el árbol filogenético.
Se encontraron genomas muy grandes, con un valor C de unos 350 picogramos, en algunas Allioideae, Amaryllidoideae, Scilloideae y Orchidaceae (Leitch et al. 2005).[12] En suma, la anatomía de la semilla parece poseer el mejor conjunto de caracteres que unifican al orden, que de otra forma es muy heterogéneo (particularmente teniendo en cuenta que Orchidaceae está dentro de él).
Asparagales comúnmente tiene micorrizas arbusculares de tipo Arum, mientras que en Liliales estas micorrizas son comúnmente de tipo Paris (ver F. A. Smith y Smith 1997).[13]
La diversidad taxonómica de las monocotiledóneas está presentada en detalle por Kubitzki (1998,[14] 2006).[15]
La familia de las orquídeas es la más grande de las angiospermas. La morfología de sus flores es muy distintiva y las hace fáciles de reconocer. La flor tiene simetría bilateral, los 3 sépalos son coloridos y vistosos (por eso algunos los llaman tépalos externos) y hay uno a cada lado y otro en el lado dorsal de la flor. Los 3 pétalos (a veces llamados tépalos internos), también vistosos, están ubicados alternadamente entre los sépalos, dos a los lados y uno en el lado ventral. Este último se llama "labelo" porque posee una morfología singular. Entre las orquídeas se encuentran algunos de los sistemas de polinización más complejos e interesantes de entre todas las angiospermas.
Las orquídeas son conocidas por poseer muchas especies de gran valor ornamental. La vainilla (de cuyo fruto se extrae la vainillina) también es una orquídea.
Borya es una planta arborescente y es típicamente una "planta reviviscente" ("resurrection plant") que se encuentra en las pendientes rocosas. Durante la estación seca estas plantas se resecan y se vuelven de un color anaranjado óxido, pero rápidamente se vuelven verdes y activas una vez empieza a llover. Junto con el otro género de boryáceas (Alania) son plantas xeromórficas nativas de Australia, la inflorescencia tiene escapo e involucro, pero las flores son difíciles de distinguir.
Blandfordia es el único género de la familia Blandfordiaceae, con 4 especies distribuidas en el este de Australia. Allí se las llama "Christmas Bells" ("campanas de Navidad") debido a la forma de sus flores y al momento de su floración, que coincide con la Navidad australiana. Son hierbas perennes erectas (de unos 1,50 m de altura), de hojas dísticas, su inflorescencia un racimo, con flores con pedicelos articulados y tépalos grandes y formando un perigonio tubular. El receptáculo floral desarrolla un ginóforo. Las anteras son latrorsas y las semillas tienen pelos conspicuos.
Lanaria lanata es la única especie de la familia Lanariaceae, distribuida en el sur de Sudáfrica. Con los caracteres más o menos típicos de las monocotiledóneas, pueden ser reconocidas por sus inflorescencias cortamente ramificadas cubiertas de pelos dendríticos, y sus flores de simetría radial con tépalos a medio connar.
Las Asteliáceas forman una familia de 2 a 4 géneros de plantas presentes en el hemisferio sur. Son más o menos rizomatosas, de hojas espiraladas, con flores en una inflorescencia que puede ser un racimo o una espiga, las brácteas de las inflorescencias son grandes, las flores pequeñas, los tépalos connados basalmente, el androceo adnato al perianto, las anteras basifijas.
Las hipoxidáceas comprenden unas 150 especies distribuidas por todo el globo salvo Europa y el norte de Asia. Pueden ser reconocidas por sus rosetas de hojas plicadas o al menos plegadas y con bases persistentes, su indumento no glandular es bastante prominente. En las flores, el verticilo externo de tépalos tiende a ser verde por fuera, y el ovario es ínfero, muchas veces hay una parte delgadamente tubular en el ápice del ovario formado por los tépalos connados o por un pico apical del mismo ovario.
Esta familia comprende un único género, Ixiolirion, con 4 especies distribuidas desde Egipto hasta el Asia Central. Son hierbas con cormos, las inflorescencias racimosas, y flores cortas tubulares azules con ovario ínfero.
Los 9 géneros de Tecofileáceas se distribuyen en Chile, Estados Unidos y África. Son hierbas con cormos, a veces con hojas de lámina ancha, pecioladas. Sus flores tienen tépalos que se expanden y tienen un androceo que las vuelve zigomorfas, ya que los estambres son fuertemente dimórficos. Las anteras se abren por poros.
El género Cyanastrum a veces es tratado en su propia familia Cyanastraceae.
Las dos especies de Doryanthes, el único género de la familia, son hierbas enormes formadoras de rosetas, son un elemento conspicuo de la flora de los alrededores de Sídney, Australia, y es difícil que su floración pase inadvertida. Las hojas tienen márgenes enteros pero se desintegran en fibras hacia el ápice. Las inflorescencias subumbeladas nacidas al final de largos tallos tienen numerosas flores de color rojo brillante, de simetría radial y con ovario ínfero.
Las iridáceas son unos 70 géneros con más de 1600 especies que se distribuyen por todo el globo.
Se diferencian de otras familias relacionadas en que son usualmente hierbas perennes con hojas generalmente ensiformes y unifaciales, la inflorescencia es una espiga o panícula bracteada de flores solitarias o agrupaciones de cimas monocasiales (ripidios), y las flores poseen solo 3 estambres, opuestos a los tépalos externos.
Dentro de esta familia se encuentra Crocus sativus L., de la cual se obtiene el azafrán a partir de sus estilos secos. Muchas especies de Iridaceae presentan una gran importancia económica en la horticultura ornamental y en la industria de la flor cortada, especialmente Gladiolus, Freesia, Sparaxis, Iris, Tigridia ("flor tigre"), Ixia ("lirio del maíz"), Romulea, Neomarica, Moraea ("lirio mariposa"), Nemastylis, Belamcanda, Sisyrinchium ("pasto de ojos azules"), Crocosmia, y Trimezia. Muchos otros géneros (Watsonia, Crocus, Dietes, Tritonia, Hesperantha y Neomarica) se cultivan en jardines en regiones tropicales y templadas, como plantas perennes y bulbosas.[16][17][18]
Moraea y Homeria son dos géneros de plantas venenosas y representan un problema en las regiones productoras de ovinos y bovinos, notablemente en Sudáfrica.[16] Los cormos de algunas especies son utilizados como alimento por algunos pueblos indígenas.
Las xeronematáceas, con su único género Xeronema, están distribuidas en la isla Poor Knights de Nueva Zelanda, y en Nueva Caledonia. Son hierbas bastante grandes que pueden ser reconocidas por sus hojas isobifaciales equitantes, y sus inflorescencias abarrotadas con flores bastante grandes, de simetría radial, que miran hacia arriba. Los estambres extán fuertemente exsertos. La familia es pobremente conocida aún.
Hemerocallidaceae incluye plantas herbáceas, glabras, perennes, provistas de cortos rizomas y raíces fibrosas o rizomatosas con tubérculos radicales. Las hojas son arrosetadas en la base de la planta, alternas, dísticas, planas, sésiles, simples, linear o lanceoladas, paralelinervadas. Los márgenes de la lámina son enteros. Las flores son leve o acusadamente cigomorfas, hermafroditas y presentan nectarios. Se hallan dispuestas en varios tipos de inflorescencias, tales como cima helicoidal y racimo, en la extremidad de un escapo bracteado. Incluye 8 géneros y aproximadamente 85 especies distribuidas en zonas templadas de Europa y Asia, Malasia, India, Madagascar, África y el Pacífico, especialmente Australia y Nueva Zelandia; hasta Sudamérica. Dos de los géneros, Hemerocallis y Phormium, se cultivan como ornamentales en todo el mundo.
Las xantorroeoideas están distribuidas por Australia. Pueden ser reconocidas por su hábito, ya que su tallo es erecto y leñoso (y con un indumento persistente de hojas secas si no hay incendios), y en su extremo posee una agregación densa de hojas largas y delgadas. La inflorescencia es de tipo espiga, erecta y densamente apretada. Las flores son pequeñas y el fruto una cápsula.
Es una subfamilia de Xanthorrhoeaceae nativa de regiones templadas a tropicales del Viejo Mundo, con 15 géneros y 780 especies. La mayor diversidad ocurre en Sudáfrica, usualmente en hábitats áridos. Se distinguen de otros clados relacionados por ser hierbas o árboles paquicaulos (esto es, con un tronco más ancho en su parte basal, la parte ancha funcionando en almacenamiento), con hojas usualmente suculentas, flores trímeras con un ovario súpero, y las semillas con arilo. El género más conspicuo de la familia es Aloe. Muchas especies de Aloe son usadas medicinalmente o en cosméticos. Por ejemplo, el "aloin" es derivado del Aloe vera y del Aloe ferox, y tiene importantes usos médicos, por ejemplo como laxantes y en tratamientos de quemaduras, como en productos para la piel, el cabello y la salud. Miembros de muchos géneros son usados como ornamentales, por ejemplo Aloe, Haworthia, Gasteria, Kniphofia, Asphodelus y Bulbine.
Agapanthus es el único género de esta subfamilia de las amarilidáceas, endémico de Sudáfrica. Son plantas herbáceas, perennes, más bien robustas, provistas de cortos rizomas y hojas arrosetadas, linear-oblongas, planas, más bien carnosas. Las flores son en general grandes, azules o blancas y se hallan dispuestas en umbelas en la extremidad de un escapo áfilo, más largo que las hojas. Las inflorescencias se hallan protegidas por brácteas soldadas entre sí a lo largo de uno de sus lados. El ovario es súpero. No presentan el olor a ajo característico de las Allioideae y los análisis moleculares de ADN demuestran que son más cercanas a las Amaryllidoideae. No obstante, se separa de este último clado por el tipo de ovario, la presencia de saponinas y la ausencia de los alcaloides típicos de las amarilidoideas.
Las alioideas se pueden encontrar generalmente en las regiones templadas, cálidas y subtropicales de todo el mundo. Son herbáceas y perennes, y normalmente poseen bulbo, aunque en algunos casos tienen rizoma. Las especies que componen la familia pueden ser reconocidas fácilmente por su olor característico (el olor de los ajos y las cebollas, suficientemente singular como para ser denominado olor aliáceo), por sus hojas bastante suaves y carnosas y su inflorescencia parecida a una umbela en la extremidad de un escapo, la cual lleva flores pequeñas a medianas de ovario súpero. Se las cultiva como hortalizas y condimenticias, como plantas medicinales y también como ornamentales. En el primer caso, el género económicamente más importante es Allium, que incluye algunas de las plantas comestibles más utilizadas, tales como la cebolla (Allium cepa), el ajo (A. sativum y también A. scordoprasum), la chalota, echalote o escalonia (Allium ascalonicum), el puerro (A. ampeloprasum), el puerro salvaje o rampa (Allium tricoccum) y varios condimentos como la cebolleta (A. schoenoprasum). Por otro lado, los compuestos organosulfurados de las aliáceas tienen propiedades antioxidantes, antibióticas, anticarcinogénicas, antiteratogénicas y estimulatorias del sistema inmune y protectoras de la función hepática.[19]
La familia también posee importantes ornamentales.
Esta familia comprende 59 géneros y aproximadamente 800 especies propias de regiones sobre todo templadas y cálidas, de todos los continentes. Son plantas herbáceas perennes con bulbo, y pueden ser fácilmente reconocidas por sus hojas bastante carnosas, sus flores generalmente grandes y atractivas, con 6 estambres y un ovario ínfero, las flores se hallan solitarias o, más frecuentemente, dispuestas en inflorescencias en umbela en la extremidad de un escapo.
Muchas de las especies de amarilidoideas son populares como plantas ornamentales en parques y jardines. Una mención aparte merece el caso de Narcissus (el narciso), cultivado en varias partes del mundo como ornamental en jardines y también para flor de corte.
Las afilantoideas comprenden una sola especie, Aphyllanthes monspeliensis, distribuida en las zonas áridas del oeste del Mediterráneo. La planta está comprendida por agrupamientos de inflorescencias escaposas, los escapos son en realidad el principal órgano fotosintético, ya que las hojas escariosas en la base no son fotosintéticas. La inflorescencia tiene pocas flores, con tépalos usualmente azules de tamaño mediano. La planta es parecida a Sisyrinchium.
Las brodiaeoideas (antes familia de las temidáceas) comprenden una docena de géneros nativos del oeste de Norteamérica. Son muy similares a las alioideas, ya que son hierbas perennes con una inflorescencia umbelada al final de un escapo, con flores bastante pequeñas, y tépalos más o menos conados a veces con una corona, y el ovario es súpero, pero carecen del "olor aliáceo" típico de estas últimas, y poseen un cormo fibroso, no un bulbo, y además hay usualmente 4 o más brácteas en las inflorescencias que no son como las 3 brácteas de las alioideas, que son más envolventes.
Las scilloideas (antes familia de las hiacintáceas) comprenden de 770 a 1000 especies, distribuidas predominantemente en climas mediterráneos, especialmente Sudáfrica y el Mediterráneo hasta Asia Central y Birmania (algunas en Sudamérica). Poseen 6 tépalos y 6 estambres con ovario súpero, por eso se las solía anidar en Liliaceae. Se caracterizan por ser herbáceas perennes con bulbo, con hojas bastante carnosas y mucilaginosas que se disponen en una roseta basal. Poseen compuestos venenosos, por lo que sus especies no son comestibles. Algunos géneros son muy populares en jardinería, como Hyacinthus (el conocido jacinto), importante como planta cultivada y como flor cortada.
Las agavoideas están distribuidas más o menos por todo el mundo y son especialmente diversas en México, donde Agave tiene importantes usos económicos (por ejemplo es utilizado para elaborar tequila y mescal). Muchos miembros de esta subfamilia son arborescentes, como el árbol de Josué (una especie de Yucca). Las hojas son muchas veces grandes, xeromórficas, fibrosas o raramente suculentas, en rosetas en la base o el final de las ramas. Las flores presentan 6 tépalos y 6 estambres, y el ovario es súpero o ínfero.
Las lomandroideas (antes familia de las laxmaniáceas) son predominantemente australianas. Poseen los tépalos persistentes en el fruto. La familia puede ser dividida en 3 grupos con caracteres distintivos: el grupo Lomandra, el grupo Cordyline y el grupo Laxmannia.
Las asparagoideas están distribuidas por el Viejo Mundo (Asparagus) y México (Hemiphylacus). Los órganos fotosintéticos aplanados de Asparagus han sido sujeto de mucha controversia, sin embargo, la mayoría de los autores los consideran tallos aplanados. Es curioso que ramas fotosintéticas similares hayan evolucionado independientemente en Nolinoideae (antes Ruscaceae), subfamilia con la que las asparagoideas están emparentadas. Asparagus posee hojas reducidas a escamas no fotosintéticas que llevan en su axila filoclados verdes, solitarios a abigarrados, circulares a aplanados en sección transversal, y el fruto es una baya.
Las nolinoideas (antes familia de las ruscáceas), como hoy definidas (en un sentido más amplio que el tradicional), están ampliamente distribuidas en regiones templadas a tropicales. Son herbáceas a arborescentes, a veces tienen crecimiento secundario anómalo, a veces los tallos son aplanados y fotosintéticos (son filoclados). Las flores bisexuales, de simetría radial, son pequeñas. Los 6 tépalos suelen estar conados en la base por lo que las flores tienen forma de campana o similares. Poseen 6 estambres y un ovario súpero. El fruto usualmente es una baya de pocas semillas.
La monofilia de Asparagales está sostenida por análisis cladísticos basados en morfología, ADNr 18S, y muchas secuencias de ADN (Chase et al. 1995a,[20]b,[21] 2000,[22] 2006,[23] Conran 1989,[24] Davis et al. 2004,[25] Graham et al. 2006,[26] Hilu et al. 2003,[27] Pires et al. 2006,[28] Rudall et al. 1997,[29] Källersjö et al. 1998,[30] Fay et al. 2000,[31] Soltis et al. 2000,[32] Stevenson et al. 2000,[33] McPherson y Graham 2001),[34] si bien algunas reconstrucciones filogenéticas como la de Hilu et al. (2003),[27] con datos moleculares, sugirieron que Asparagales puede ser parafilético, con Orchidaceae separado del resto.
Con respecto a las relaciones de Asparagales con el resto de los órdenes de monocotiledóneas, es el clado hermano del clado Commelinidae.
Han hecho análisis morfológicos Chase et al. (1995b)[21] y Stevenson y Loconte (1995),[35] y en los dos al menos algunas de las familias de Asparagales con crecimiento secundario anómalo formaban clados, si bien también caían dentro de estos clados algunos miembros que hoy están ubicados en Liliales, Zingiberales y Pandanales. También ver Rudall (2002)[36] para un trabajo reciente de filogenia del orden con datos morfológicos.
Las relaciones dentro del orden fueron investigadas por Dahlgren et al. (1985),[6] Rudall y Cutler (1995),[37] Chase et al. (1995a,[20]b,[21] 1996),[38] Stevenson y Loconte (1995),[35] Rudall et al. (1997),[29] Fay et al. (2000),[31] McPherson y Graham (2001),[34] Graham et al. (2006)[26] y Pires et al. (2006).[28]
El cladograma resumido, según Judd et al. 2007 (modificado de Soltis et al. 2005,[39] Orchidaceae basal como en APWeb 2008):[4]
AsparagalesHypoxidaceae (tradicionalmente ubicado en Amaryllidaceae)
Xanthorrhoeaceae sensu latoHemerocallidoideae (tradicionalmente ubicado en Liliaceae)
Xanthorrhoeoideae (tradicionalmente ubicado en Liliaceae)
Asphodeloideae (tradicionalmente ubicado en Liliaceae)
Altos Asparagales Amaryllidaceae sensu latoAgapanthoideae (tradicionalmente ubicado en Liliaceae)
Allioideae (tradicionalmente ubicado en Liliaceae)
Lomandroideae (antes familia Laxmanniaceae, tradicionalmente ubicado en Liliaceae)
Asparagoideae (tradicionalmente ubicado en Liliaceae)
Nolinolidease (antes familia Ruscaceae, tradicionalmente ubicado en Liliaceae)
Brodiaeoideae (antes familia Themidaceae, tradicionalmente ubicado en Liliaceae)
Scilloideae (antes Hyacinthaceae, tradicionalmente ubicado en Liliaceae)
Agavoideae (tradicionalmente ubicado en Amaryllidaceae o Liliaceae)
Un cladograma completo es proporcionado por el APWeb.[4]
Asparagales ¿Hypoxidaceae sensu lato? Xanthorrhoeaceae sensu lato Altos Asparagales (= Núcleo de los Asparagales) Amaryllidaceae sensu lato Asparagaceae sensu latoBrodiaeoideae = Themidaceae
Scilloideae = Hyacinthaceae
Agavoideae (Anemarrhena, Agave y afines, Hesperocallis, Behnia, Herreria y afines, Anthericum y afines)
Lomandroideae = Laxmanniaceae
Nolinoideae = Ruscaceae
El árbol, con los nombres de los clados actualizados según el APG III (2009),[1] está basado principalmente en los análisis de Chase et al. (2000)[22] y Fay et al. (2000).[31] Estos estudios difieren poco en los detalles, si bien el análisis de Fay et al. (2000)[31] tiene más nodos con buen apoyo en el núcleo de los Asparagales (lo cual no es sorprendente, ya que utilizaron el método de pesaje sucesivo o "successive weighting"). También es largamente congruente con esta filogenia la presentada por McPherson y Graham (2001),[34] si bien su muestreo es mucho menor.
De los análisis se desprende un asemblaje basal parafilético, llamado los "Bajos Asparagales" (sensu Rudall et al. 1997),[29] que incluye a familias como Orchidaceae, Hypoxidaceae, Iridaceae, Asphodeloideae, y Hemerocallidoideae. En contraste, los llamados "Altos Asparagales" son un clado propiamente dicho que se desprende del resto, e incluye a Allioideae, Amaryllidoideae, Scillloideae = Hyacinthaceae, Agavoideae, Asparagoideae, y Nolinoideae = Ruscaceae. Muchos de los "Bajos Asparagales" se caracterizan por la microsporogénesis simultánea (las 4 microsporas se separan a un tiempo entre sí después de las dos divisiones meióticas). El patrón de microsporogénesis de los "Bajos Asparagales" es apomórfico dentro de las monocotiledóneas. El patrón de microsporogénesis de los "Altos Asparagales" en cambio, es como aquellos de las monocotiledóneas más basales. Rudall (2001,[40] véase también 2002b,[41] 2003a)[42] incluyó al carácter del ovario ínfero como una sinapomorfía del orden, notando que en los "Altos Asparagales" podría haber habido una reversión al ovario súpero, reversión asociada con la presencia de nectarios septales infraloculares (como en Xanthorrhoea y Johnsonia, dos géneros de Hemerocallidoideae). Sin embargo, Stevens (2001 en adelante, APWeb)[4] nota que los ovarios súperos también aparecen distribuidos todo a lo largo de los "Bajos Asparagales", por lo que dónde deben ubicarse en el árbol las evoluciones de las diferentes morfologías del ovario no está claro: la posición del ovario parece un carácter mucho más flexible (aquí como en las demás angiospermas) de lo que se creía. Los "Altos Asparagales", por ser un grupo monofilético con fuerte apoyo en los análisis moleculares de ADN, son llamados "Núcleo de los Asparagales" ("Core Asparagales") por el APWeb.[4]
Orchidaceae es la familia más grande de todas las angiospermas. Algunos autores (por ejemplo Dahlgren et al. 1985)[6] reconocieron tres familias de orquídeas, pero los análisis moleculares de ADN demostraron que estas familias son polifiléticas y deberían reunirse en un Orchidaceae sensu lato, que es como se trata a la familia hoy en día. Con respecto a su relación con las demás familias de Asparagales, varios trabajos recientes sugieren que Orchidaceae es hermana del resto de los Asparagales (por ejemplo con apoyo bootstrap del 76 % en Graham et al. 2006,[26] un resultado más o menos similar en Givnish et al. 2006b,[43] un apoyo más fuerte —del 96 al 99 %—, con un buen tamaño de muestreo y 7 genes de dos compartimentos, en Pires et al. 2006,[28] y también un buen apoyo en Chase et al. 2005,[44] donde el nodo separando a las orquídeas del resto de Asparagales tenía 91 % de apoyo bootstrap). Otros trabajos la ubicaron en forma diferente en el árbol filogenético: a veces fue ubicada como hermana de Boryaceae (por ejemplo en Chase et al. 1995a[20] y en McPherson y Graham 2001,[34] si bien con un apoyo más bien bajo). En cambio Li y Zhou (2007)[45] encuentran a Orchidaceae embebida en un clado parafilético Boryaceae-Hypoxidaceae. Por otro lado Rudall (2003a),[42] con datos morfológicos, sugiere una relación morfológica cercana entre Hypoxidaceae y Orchidaceae en particular. Janssen y Bremer (2004),[46] si bien no ubican a Orchidales como hermanos del resto del orden, lo ubican temporalmente cerca del comienzo de la divergencia dentro de él (aunque la topología del árbol que utilizaron difiere considerablemente en detalle del utilizado aquí). En resumen, la posición de Orchidaceae como hermano del resto de Asparagales, con Boryaceae como parte del clado que incluye a Blandfordiaceae, parece la mejor hipótesis, y esta topología es la presentada aquí. Nótese que esto cambia la caracterización de Asparagales, muchos de sus caracteres ahora habiendo sido reubicados al nodo subbasal en el clado (comparar la lista de apomorfías del APWeb[4] en versiones 7 y anteriores a esa del sitio web). Un dato interesante es que las orquídeas carecen de la mayoría de las sinapomorfías no moleculares del orden: las semillas de Orchidaceae no tienen fitomelaninas debido a su naturaleza (son como un polvillo, como la mayoría de los micoparásitos), y sus nectarios raramente están en los septos de los ovarios. Las orquídeas tienen microsporogénesis simultánea y ovario ínfero, caracteres que son típicos de los "Bajos Asparagales" de Rudall et al. 1997,[29] pero no está claro si estos caracteres deberían considerarse sinapomorfías del orden. La única sinapomorfía de Orchidaceae aparentemente es la presencia de protocormo (la estructura producida por el crecimiento de sus embriones indiferenciados antes de que las raíces y tallos se desarrollen). Las orquídeas presentan el androceo y el gineceo fusionados, pero esta característica está presente solo en la subfamilia Apostasioideae y solo en grados variables. Las semillas como polvillo dispersadas por viento están presentes en la mayoría de las subfamilias, pero en algunos miembros de Cypripedioideae y Vanilloideae hay semillas crustosas (probablemente asociadas a su forma de dispersión, que en este caso es por pájaros y mamíferos que son atraídos por su fruto carnoso que fermenta in situ liberando compuestos fragantes, por ejemplo la vainillina). La diversificación en Orchidaceae es remarcable, pero debe ser comparada con la de los demás Asparagales, que son menos ricos en especies pero muy variables morfológicamente.
Las relaciones de Boryaceae (que comprende solo dos géneros, Borya y Alania) con el resto de las familias de Asparagales permanecieron poco claras durante mucho tiempo. Las boryáceas son micorrícicas, pero no como las orquídeas, sino que sus micorrizas son del tipo estandarizado vesicular-arbuscular (VA). Previamente se llegó a pensar que Boryaceae era un miembro de Anthericaceae (Dahlgren et al. 1985,[6] Takhtajan 1997),[47] una familia que demostró ser extremadamente polifilética (Chase et al. 1996).[38] La ubicación de Orchidaceae como hermana de todo el resto de Asparagales le da un apoyo más o menos importante a la ubicación de Boryaceae como hermano del clado que incluye a Blandfordiaceae. Rudall (2003a)[42] también sugirió una relación morfológica cercana entre Boryaceae y Blandfordiaceae. Sin embargo hay que tener en cuenta que si bien hay buen apoyo en el árbol de Chase et al. (2006)[23] para posicionar a Orchidaceae como hermano de todas las demás Asparagales, Boryaceae está ubicado inmediatamente por encima del clado que incluye a Blandfordiaceae, si bien con muy bajo apoyo.
Hypoxidaceae es una familia que tradicionalmente había sido ubicada en Amaryllidoideae. Lanariaceae con su único género Lanaria en el pasado fue considerado un miembro de Haemodoraceae (Hutchinson 1967)[48] o Tecophilaeaceae (Dahlgren et al. 1985).[6] Estas dos familias junto con Asteliaceae, Blandfordiaceae y probablemente también Boryaceae forman un clado, y salvo por la ubicación de Boryaceae, las otras 4 tienen buen apoyo como clado en los análisis moleculares. Una sinapomorfía potencial para estas familias es la estructura del óvulo: al menos Asteliaceae, Blandfordiaceae, Lanariaceae e Hypoxidaceae poseen una constricción chalazal y una cofia nuclear ("nucellar cap"). Soltis et al. (2005), dejando afuera del clado a Boryaceae, sugirieron que estas familias se fusionararan en un Hypoxidaceae sensu lato.
Dentro de este clado, Asteliaceae + Hypoxidaceae + Lanariaceae forman a su vez un clado con apoyo del 100 % de bootstrap. La morfología también provee algún apoyo para estas relaciones: Asteliaceae e Hypoxidaceae forman rosetas cubiertas con pelos multicelulares ramificados (también Lanariaceae posee pelos multicelulares), y poseen canales radiculares llenos de mucílago (este último carácter también presente en Lanariaceae), pero Blandfordia (único género de Blandfordiaceae) no comparte estos caracteres. Además, este clado posee estomas paracíticos, la lámina de la hoja con una vena media distintiva, quizás el gineceo más o menos ínfero, y una micropila biestomal. Asteliaceae + Hypoxidaceae poseen flavonoles, endosperma de paredes delgadas, cotiledón no fotosintético, y la lígula larga.
El siguiente nodo en Asparagales (todas las familias menos Orchidaceae y el clado mencionado) se ha demostrado que es robusto (81 % de apoyo jacknife en Soltis et al. 2000,[32] 100 % de apoyo bootstrap en Chase et al. 2005,[44] véase también Fay et al. 2000[31] usando datos de cuatro genes plastídicos, y Chase et al. 2000[22]), pero no hay caracteres fuera de los moleculares que unan a este clado.
La posición de Ixioliriaceae y Tecophilaeaceae todavía es poco clara. Algunos análisis moleculares las unen en un clado, por ejemplo recibe apoyo de leve a moderado en Chase et al. (2000),[22] Pires et al. (2006),[28] y Givnish et al. (2006),[43] y en Graham et al. (2006)[26] recibe un apoyo más fuerte pero el muestreo es más bien pobre. En el análisis de 3 genes de Fay et al. (2000)[31] el clado tiene un brazo muy largo. Estas dos familias comparten los cormos, la inflorescencia foliosa, y muchas veces, una inflorescencia casi capitada, las hojas de disposición espiralada y base envainadora, las flores más bien grandes, el verticilo externo de tépalos mucronado a aristado, el perianto formando un tubo corto, el androceo insertado en la boca, y un número cromosómico x=12 (Soltis et al. 2005 y APWeb). También se encuentran otras topologías: en Janssen y Bremer (2004)[46] Ixioliriaceae diverge considerablemente antes (si bien es adyacente a) Tecophilaeaceae; y en Davis et al. (2004),[25] con un muestreo pobre, se encuentra algún apoyo para hermanar a Ixioliriaceae con Iridaceae, también en Chase et al. (2006)[23] se encuentra apoyo (esta vez fuerte) para este parentesco, en donde el clado es hermano de Doryanthaceae (pero con apoyo bajo).
Las relaciones de Doryanthaceae con las familias que la rodean en los análisis filogenéticos todavía son poco claras tanto en los análisis morfológicos como en los moleculares. Chase et al. (2006)[23] la encontró en la ubicación recién mencionada, que no es la presentada aquí. Para la ubicación presentada aquí hay apoyo moderado en Fay et al. (2000),[31] y un apoyo de 92 % en Graham et al. (2006).[26] Rudall (2003)[42] encuentra una relación morfológica cercana entre Doryanthaceae e Iridaceae.
El nodo que contiene a Iridaceae y a todo el resto de los Asparagales (Xeronemataceae, etc.) recibe un apoyo bastante fuerte en Fay et al. (2000).[31] El grupo tiene algunas características moleculares en común: ha perdido los telómeros de tipo Arabidopsis, mientras que son comunes los telómeros de tipo humano (TTAGGG).
Iridaceae es una de las familias más grandes y mejor estudiadas de Asparagales (por ejemplo ver Goldblatt 1990,[49] 1991,[50] Goldblatt et al. 1998,[51] 2002,[52] Reeves et al. 2001).[53] Iridaceae es distintiva entre los Asparagales debido a la estructura única de su inflorescencia (un ripidio) y su combinación de ovario ínfero con 3 estambres, también son comunes en la familia las hojas unifaciales, mientras que las hojas bifaciales son la norma en otros Asparagales.
El nodo que sigue al posible desprendimiento de Iridaceae está bien sostenido en los análisis (Fay et al. 2000,[31] Chase et al. 2005).[44] Si un ovario ínfero es una sinapomorfía de Asparagales (Chase et al. 1995b)[21] entonces este nodo marcaría la transición a un ovario súpero (aunque se encuentran ovarios ínferos en Amaryllidoideae y en Yucca). Todos los clados de este nodo tienen nectarios septales infraloculares, lo que Rudall (2000)[54] interpreta como ovarios secundariamente súperos. Esta interpretación demuestra la reversibilidad de este carácter que muchas veces fue enfatizado en la sistemática de monocotiledóneas (por ejemplo en Cronquist 1981,[55] Dahlgren et al. 1985).[6] Es en este nodo en el que se pierde el gen mitocondrial rpl2 (o quizás en el siguiente, de acuerdo con la distribución en Adams et al. 2002).[56]
Xeronemataceae con su único género Xeronema fue previamente considerado cercanamente emparentado con Phormium (usualmente Phormiaceae, Dahlgren et al. 1985),[6] pero si bien hay similitudes superficiales (como las hojas unifaciales), no son idénticos en los detalles. Chase et al. (2000)[22] recientemente describió a Xeronemataceae, y sus caracteres varían según patrones que son generales en Asparagales.
El siguiente nodo del árbol (Xanthorrhoeaceae sensu lato + los "Altos Asparagales") tiene fuerte apoyo en Fay et al. (2000)[31] y en Chase et al. (2000c).[57]
Hemerocallidoideae está cercana a Asphodelaceoideae y Xanthorrhoeoideae, y los 3 clados tienen antraquinonas, estiloides, y semilla con cotiledón no fotosintético. Ver Kite et al. (2000)[58] para la distribución de antraquinonas, McPherson et al. (2004)[59] para los taxones que no tienen el intrón 3'-rps12. Dentro de este clado, ocurre crecimiento secundario (anómalo) en Aloe (algunas especies llegando a tamaños enormes), Phormium y Xanthorrhoea. Las 3 familias son por algunos autores reunidas en un Xanthorrhoeaceae sensu lato (como en APG III y en el APWeb, 2001 en adelante),[4] aunque Stevens en el APWeb remarca que la microsporogénesis es variable en este grupo. En Fay et al. (2000)[31] se encuentra fuerte apoyo para este clado. Sin embargo, las relaciones de parentesco dentro de él aún no están esclarecidas. En el análisis de 3 genes de Chase et al. (2000)[22] hay un apoyo moderado para la relación (Xanthorrhoeoideae + Asphodeloideae), que entre los caracteres morfológicos encuentra la presencia de engrosamiento secundario, el androceo no adnato a los tépalos, la presencia de hipostasa ("hypostase"), y las semillas angulosas. Esta relación es la presentada aquí, sin embargo el apoyo disminuye cuando se incluyen los taxones donde faltan algunas secuencias de genes, véase también Fay et al. 2000).[31] Sin embargo, Devey et al. (2006)[60] encuentran algún apoyo para reunir a Xanthorrhoeoideae con Hemerocallidoideae (véase también Pires et al. 2006),[28] mientras que Chase et al. (2006)[23] sugieren un clado formado por Asphodeloideae + Hemerocallidoideae (pero el muestreo es cuestionado en el APWeb).[4] Con respecto a la morfología, tanto Hemerocallidoideae como Xanthorrhoeoideae tienen ovarios que probablemente son secundariamente súperos, y tienen nectarios septales infraloculares (Rudall 2002,[36] 2003).[42] Rudall (2003)[42] sugiere una relación morfológica cercana entre Hemerocallidoideae y Asphodeloideae, y otra entre Xanthorrhoeoideae e Iridaceae.
El valor de las antraquinonas como carácter taxonómico es apreciado ahora, ya que Asphodeloideae las posee pero anteriormente no había sido ubicada como cercana a las otras dos familias de Xanthorrhoeaceae sensu lato sino a Anthericaceae (con la cual ahora se sabe que está lejanamente relacionada). Fue debido a la presencia de antraquinonas que Asphodeloideae fue ubicada separada de Anthericaceae por Dahlgren et al. (1985),[6] además de que Asphodeloideae tiene microsporogénesis simultánea mientras que Anthericaceae tiene microsporogénesis sucesiva. Asphodeloideae tiene muchos géneros con cariotipos bimodales distintivos (Chase et al. 2000c),[57] y este carácter fue usado de base para sugerir que estos géneros fueran separados en una familia Aloaceae (Dahlgren et al. 1985).[6] Sin embargo los estudios moleculares mantuvieron que Asphodeloideae se quedara en su circunscripción amplia.
Xanthorrhoeoideae antes incluía otros taxones arborescentes como Kingia (que no tiene crecimiento secundario y ahora es ubicado en Dasypogonaceae) y Cordyline (con el cual está solo lejanamente emparentado, es un miembro de Lomandroideae, antes Laxmanniaceae).
Hemerocallidaceae muchas veces comprendía solo al género Hemerocallis (Dahlgren et al. 1985),[6] pero ya desde la circunscripción del APG (1998),[7] la familia incluía un gran número de géneros, como Dianella, Johnsonia, y Phormium. Todos estos géneros tienen polen tricotomosulcado, una rareza entre las monocotiledóneas, y los estudios moleculares tempranos (Chase et al. 1995a)[20] fueron críticos en determinar si este carácter era importante para estos géneros (Rudall et al. 1997).[29] En el APG III es la subfamilia Hemerocallidoideae de Xanthorrhoeaceae sensu lato.
El resto de las familias, que se podrían reunir en los clados Amaryllidaceae sensu lato y Asparagaceae sensu lato, se denomina en conjunto "Núcleo de los Asparagales" por el APWeb,[4] se caracteriza por poseer microsporogénesis sucesiva (son los "Altos Asparagoides" de Rudall et al. 1997),[29] y quizás también por poseer saponinas esteroideas. En los análisis moleculares de ADN es un grupo con fuerte apoyo (por ejemplo en Chase et al. 1995a,[20] Fay et al. 2000,[31] Chase et al. 2000c,[57] Graham et al. 2005).[61] Sin embargo, la inclusión de Aphyllanthoideae (en Asparagaceae sensu lato), tiende a disminuir el apoyo (Graham et al. 2006).[26] Además, hay poco apoyo morfológico especialmente para las agrupaciones dentro del área de Asparagaceae.
Amaryllidaceae sensu lato (Allioideae, Amaryllidoideae y Agapanthoideae) está reunido por sus secuencias de ADNcp (Fay et al. 2000,[31] pero ver McPherson et al. 2004[59] y Thomas et al. 2005) y sus secuencias ADNnr ITS para Amaryllidoideae + Agapanthoideae (Meerow et al. 2000.[62] Dentro del clado, [Agapanthoideae [Allioideae + Amaryllidoideae]] es sugerido por Meerow et al. (1999),[63] Fay et al. (2000,[31] con fuerte apoyo), Givnish et al. (2006)[43] y Pires et al. (2006),[28] pero Meerow et al. (2000a)[64] encontraron a Agapanthoideae como hermana de Amaryllidoideae, si bien con bajo apoyo.
El clado posee algunos caracteres morfológicos: las inflorescencias son escaposas, umbeloides (construcción cimosa), generalmente con dos (o más) brácteas como espatas escariosas fusionadas encerrándolas. Las hojas son dísticas, los pedicelos no articulados, el estilo largo, el estigma seco, el endosperma nuclear o helobial, sin hipocótilo, quizás se debiera incluir a los tépalos libres, las anteras conadas en la base, y las células del tapete uninucleadas. (Allioideae + Amaryllidoideae) posee como posibles sinapomorfías el hecho de que son geofitas, el tallo que es un bulbo de ramificación simpodial, tunicado, y las raíces contráctiles.
Las 7 familias que forman el clado Asparagaceae sensu lato están bien sostenidas en el análisis de Fay et al. (2000,[31] sin incluir Hesperocallis), pero no hay caracteres morfológicos muy evidentes que reúnan al clado, aunque es buen candidato el endosperma: helobial, de paredes gruesas, hemicelulósico. Incluso las familias en sentido estricto de este clado son difíciles de reconocer, porque no poseen sinapomorfías obvias, y sus flores tienen el típico aspecto de las liliáceas (en ese sentido el APWeb[4] sugiere que reunirlas en un Asparagaceae sensu lato podría ser útil, como de hecho el APG III en el 2009 lo hace). Para detalles de sus relaciones, ver Jang y Pfosser (2002)[65] y Bogler et al. (2006).[66]
En Asparagaceae sensu lato, a diferencia de Amaryllidaceae sensu lato, las inflorescencias son racimos (si bien Brodiaeoideae = Themidaceae tiene inflorescencias umbeloides pero en este caso, tienen brácteas por debajo de todas las flores, por lo que las flores se interpretan como racimos condensados). En Asparagaceae sensu lato hay muchos géneros arborescentes (como Yucca, Agave, Cordyline), y también algunos con raíces con crecimiento secundario (como Herreria, Thysanotus), y muchos son geofitas. Amaryllidaceae sensu lato es mucho menos diversa, con la mayoría geofitas, bulbosas, si bien algunas son rizomatosas (como Agapanthus, Tulbaghia, Clivia). Un carácter morfológico que reúne a las asparagáceas sensu lato es la presencia de una inflorescencia racimosa.
Brodiaeoideae = Themidaceae, Scilloideae = Hyacinthaceae, Aphyllanthoideae y Agavoideae forman un clado (ver Fay et al. 2000[31] para todas menos Aphyllanthoideae), si bien el sostén morfológico para este grupo es poco claro. Este clado puede ser hermano del clado (Nolinoideae = Ruscaceae + Asparagoideae + Lomandroideae = Laxmanniaceae).
Las relaciones entre Aphyllanthoideae, Brodiaeoideae = Themidaceae y Scilloideae = Hyacinthaceae quizás deban ser representadas como una tricotomía. Fay et al. (2000),[31] Pires et al. (2001),[67] y Pires y Sytsma (2002)[68] discuten la incertidumbre con respecto a las relaciones de Brodiaeoideae. (Brodiaeoideae + Scilloideae) parecen tener un moderado sostén en Fay y Chase (1996)[69] y en Meerow et al. (2000),[64] pero el sostén es bajo en el análisis de 2 genes de Jang y Pfosser (2002,[65] sin incluir Aphyllanthes), en Chase et al. (2006),[23] y en Pires et al. (2006).[28] Los dos clados comparten las saponinas esteroideas, las hojas de disposición espiralada, los pedicelos bracteados, el óvulo anátropo, el endosperma helobial o nuclear, y el cotiledón no fotosintético. Para algunos caracteres más ver Fay y Chase (1996).[69] Estructuras parecidas a laticíferos pueden estar presentes en los dos clados.
Aphyllanthes tiene una rama muy larga en el análisis de 3 genes de Fay et al. (2000),[31] y su posición filogenética es poco clara, pero su remoción de algunos análisis disminuye drásticamente el apoyo (Chase et al. 2006).[23] Una relación de Aphyllantes como hermano de Scilloideae fue encontrada por McPherson y Graham (2001),[34] pero Pires et al. (2006)[28] lo encontraron como hermano de Lomandroideae, pero con apoyo bajo.
Las subfamilias Lomandroideae + Asparagoideae + Nolinoideae comparten las saponinas esteroideas, los pedicelos articulados, el fruto que es una cápsula, el endosperma helobial, de paredes gruesas, con puntuaciones, hemicelulósico. Hay apoyo moderado para este clado en el árbol de 4 genes del cloroplasto de Fay et al. (2000).[31]
Finalmente, Nolinoideae y Asparagoideae pueden estar relacionadas, como se evidencia por sus frutos indehiscentes, sus flores bastante pequeñas y su número cromosómico x=10, esta relación recibe prelilminar apoyo por algunos análisis de secuencias de ADN (Chase et al. 1995a,[20]b,[21] 1996,[38] 2000,[22] Fay et al. 2000[31]). Los frutos carnosos con semillas sin fitomelaninas son comunes en este clado, pero no se conoce si este carácter constituye una apomorfía. Poseen frutos capsulares Hemiphylacus (hermano del resto de Asparagoideae) y el clado (Comosperma + Eriospermum), que es hermano del resto de Nolinoideae, por lo que puede ser que el fruto carnoso sea derivado dentro del clado.
Para el desarrollo de los filoclados en Asparagus, Ruscus, y afines, ver Cooney-Sovetts y Sattler (1987).[70]
El sistema APG de 1998[7] y el APG II del 2003[8] reconocieron a Hesperocallidaceae con su único género Hesperocallis, pero fue ubicado en Agavaceae por los últimos análisis moleculares de ADN (Pires et al. 2004),[71] por eso la familia fue incluida en Agavaceae en APG III del 2009[1] y en el APWeb.[4]
El orden fue reconocido por el APG III (2009),[1] el Linear APG III (2009)[2] le asignó los números de familia 62 a 75. El orden ya había sido reconocido por el APG II (2003).[8]
El orden, como actualmente está circunscripto, consiste en 14 familias y aproximadamente 26.800 especies. Los límites de algunas familias todavía son poco claros, y como se ha descripto en la sección de filogenia, algunos botánicos han propuesto que se combinen muchas de las familias que aquí se reconocen. Por ejemplo, el APG III y el APWeb tratan a Alliaceae, Amaryllidaceae y Agapanthaceae, que juntos forman un clado, como una única familia: Amaryllidaceae sensu lato. También combinan Xanthorrhoeaceae, Asphodelaceae y Hemerocallidaceae en un Xanthorrhoeaceae sensu lato, y a Agavaceae, Asparagoideae, Hyacinthaceae, Ruscaceae, Themidaceae, y unas pocas familias pequeñas sean combinadas, con la familia resultante llamada Asparagaceae sensu lato. Esta decisión es taxonómicamente arbitraria, si bien factores como la facilidad del reconocimiento y el nivel de sostén de la monofilia del grupo deberían ser considerados.
Las familias o subfamilias más representadas son Orchidaceae, Hypoxidaceae, Iridaceae, Amaryllidoideae, Allioideae, Scilloideae, Lomandroideae, Agavoideae, Asparagoideae, Convallariaceae, Asphodeloideae, y Hemerocallidoideae.
La lista de familias según APG III (2009),[1] junto con el número de familia según el Linear APG III (2009),[2] y algunas de las familias tradicionalmente reconocidas y hoy abandonadas, con su eventual destino, se listan a continuación:
Sinónimos (APWeb):[4] Asparagineae J. Presl, Asphodelineae Thorne & Reveal, Hyacinthineae Link, Iridineae Engler - Agavales Hutchinson, Alliales Traub, Amaryllidales Bromhead, Asphodelales Doweld, Asteliales Dumortier, Hypoxidales Reveal & Doweld, Iridales Rafinesque, Ixiales Lindley, Narcissales Dumortier, Orchidales Rafinesque, Tecophilaeales Reveal, Xanthorrhoeales Reveal & Doweld - Iridanae Doweld, Orchidanae Doweld - Orchididae Heintze - Crinopsida Horaninov, Orchidopsida Bartling
En el Angiosperm Phylogeny Website (julio de 2008)[4] las familias son:
El sistema APG II del 2003,[8] dispone al Orden Asparagales en el clado de las Monocotiledóneas y utiliza la siguiente circunscripción:
Nota: "+..." = Familia que opcionalmente puede segregarse de la familia precedente. O sea que, bajo este sistema un taxónomo puede, por ejemplo, ubicar al género Hemerocallis en la familia Hemerocallidaceae o bien, en la familia Xanthorrhoeaceae sensu lato (en sentido amplio).
Hesperocallidaceae con su único género Hesperocallis fue ubicado en Agavaceae por los últimos análisis moleculares de ADN, por eso fue incluida en Agavaceae en APWeb.[4]
El Sistema APG de 1998[7] también dispuso al orden Asparagales dentro del clado de las monocotiledóneas, pero utilizó una circunscripción diferente de las familias.
En el volumen 1998 (a,[14]b[72]) del Sistema Kubitzki se utilizó la siguiente circunscripción de familias dentro de Asparagales:
En el Sistema Dahlgren (1985)[6] el orden Asparagales fue dispuesto en el Superorden Lilianae dentro de la Subclase Liliidae (= Monocotiledóneas) de la Clase Magnoliopsida (= Angiospermas), y utilizó la siguiente circunscripción:
El sistema de Cronquist (1981)[55] no reconoce al orden Asparagales y dispone a muchos de sus taxones dentro del orden Liliales (en la subclase Liliidae, dentro de la clase Monocotyledoneae). Algunos géneros estaban incluso dispuestos dentro de la familia Liliaceae.
El Sistema Wettstein, que data de 1935, tampoco reconocía al orden Asparagales, y disponía a muchas de sus especies dentro del orden Liliflorales en la clase de las monocotiledóneas.
El grupo troncal Asparagales data de unos 122 millones de años hasta el presente, el grupo corona Asparagales de unos 119 millones de años hasta el presente (Janssen y Bremer 2004),[46] si bien Wikström et al. (2001)[73] sugirieron edades de 107-98 y 101-94 millones de años hasta el presente respectivamente. Hay que tener en cuenta que en el último estudio se utilizó una topología de Asparagales que difiere sustancialmente de la aceptada aquí (especialmente cerca de la base del árbol).
La divergencia del clado que contiene a todas las familias salvo Hypoxidaceae sensu lato, Boryaceae y Orchidaceae, ha sido datado en unos 84 millones de años hasta el presente (Eguiarte 1995).[74]
El "Núcleo de los Asparagales" se separa de Xanthorrhoeaceae sensu lato hace unos 93 millones de años, la divergencia dentro del clado comienza hace unos 91 millones de años (Janssen y Bremer 2004),[46] en cambio en Wikström et al. (2001)[73] se estiman fechas de 61-54 y 58-51 millones de años hasta el presente respectivamente.
El grupo troncal Alliaceae sensu lato está datado en unos 91 millones de años hasta el presente, la divergencia dentro del grupo corona Alliaceae sensu lato empieza hace unos 87 millones de años (Janssen y Bremer 2004).[46] Nótese que los hongos que infectan a Allium y otras aliáceas son bastante diferentes de los que se encuentran en Amaryllidaceae (ver por ejemplo Savile 1962).[75]
El grupo troncal Asparagaceae sensu lato está datado en unos 91 millones de años, la divergencia dentro del grupo corona Asparagaceae sensu lato empieza hace unos 89 millones de años (Janssen y Bremer 2004).[46] Eguiarte (1995),[74] sin embargo, sugiere que Agavaceae-Nolinaceae divergieron solo hace unos 47 millones de años.
|coautores= (ayuda) |fechaacceso= requiere |url= (ayuda) |coautores= (ayuda) |fechaacceso= requiere |url= (ayuda) |coautores= (ayuda) [Aliso 22: 197-203.] |coautores= (ayuda); |fechaacceso= requiere |url= (ayuda) |coautores= (ayuda); |fechaacceso= requiere |url= (ayuda) |coautores= (ayuda); |fechaacceso= requiere |url= (ayuda) |coautores= (ayuda); |fechaacceso= requiere |url= (ayuda) |coautores= (ayuda) |coautores= (ayuda) |coautores= (ayuda) |coautores= (ayuda) |coautores= (ayuda) |coautores= (ayuda) |coautores= (ayuda) |coautores= (ayuda) |coautores= (ayuda) |coautores= (ayuda) |coautores= (ayuda) |coautores= (ayuda) |coautores= (ayuda) |coautores= (ayuda); |fechaacceso= requiere |url= (ayuda) |coautores= (ayuda) |coautores= (ayuda) |coautores= (ayuda); |fechaacceso= requiere |url= (ayuda) |coautores= (ayuda) |fechaacceso= requiere |url= (ayuda) |fechaacceso= requiere |url= (ayuda) |coautores= (ayuda) [Aliso 22: 28-51.] |coautores= (ayuda) |coautores= (ayuda); |fechaacceso= requiere |url= (ayuda) (en chino) |coautores= (ayuda); |fechaacceso= requiere |url= (ayuda) |fechaacceso= requiere |url= (ayuda) |fechaacceso= requiere |url= (ayuda) |coautores= (ayuda) |coautores= (ayuda); |fechaacceso= requiere |url= (ayuda) |coautores= (ayuda); |fechaacceso= requiere |url= (ayuda) |coautores= (ayuda); |fechaacceso= requiere |url= (ayuda) |coautores= (ayuda); |fechaacceso= requiere |url= (ayuda) |coautores= (ayuda) |coautores= (ayuda); |fechaacceso= requiere |url= (ayuda) |coautores= (ayuda) [Aliso 22: 345-351.] |coautores= (ayuda); |fechaacceso= requiere |url= (ayuda) |coautores= (ayuda); |fechaacceso= requiere |url= (ayuda) |coautores= (ayuda); |fechaacceso= requiere |url= (ayuda) |coautores= (ayuda) |coautores= (ayuda); |fechaacceso= requiere |url= (ayuda) |coautores= (ayuda) [Aliso 22: 313-328.] |coautores= (ayuda); |fechaacceso= requiere |url= (ayuda) |coautores= (ayuda); |fechaacceso= requiere |url= (ayuda) |coautores= (ayuda); |fechaacceso= requiere |url= (ayuda) |coautores= (ayuda); |fechaacceso= requiere |url= (ayuda) |coautores= (ayuda); |fechaacceso= requiere |url= (ayuda) |coautores= (ayuda); |fechaacceso= requiere |url= (ayuda) |fechaacceso= requiere |url= (ayuda) |coautores= (ayuda); |fechaacceso= requiere |url= (ayuda) |coautores= (ayuda)
|coautores= (ayuda)
Asparagales es un taxón de plantas perteneciente a la categoría taxonómica de orden, utilizado en sistemas de clasificación modernos como el sistema de clasificación APG III del 2009 y el APWeb, y está circunscripto obligadamente al menos por la familia Asparagaceae. Asparagales es un orden solo recientemente reconocido en los sistemas de clasificación, y muchas de sus familias pertenecían al antiguo orden Liliales. Liliales había sido un orden muy amplio donde prácticamente se ubicaban todas las monocotiledóneas de tépalos vistosos y sin almidón en el endosperma, que había sido difícil de dividir en familias porque sus caracteres morfológicos no se daban en patrones que delimitaran grupos claramente. Con el tiempo se empezó a reconocer el orden Asparagales, especialmente cuando los análisis moleculares de ADN indicaron que Liliales debía ser dividido al menos en Liliales, Asparagales y Dioscoreales. Asparagales es un orden bastante grande, consiste en 14 familias y unas 26 000 especies. El orden está claramente circunscripto sobre la base de los análisis de secuencias de ADN pero es dificultoso de definir morfológicamente, ya que es uno de los grupos de monocotiledóneas estructuralmente más diverso. Así, el orden incluye hierbas de no más de 15 cm hasta algunas especies de Agave que superan los 10 metros de altura. En este sentido, su separación del orden Liliales es particularmente problemática.
Una de las características que definen a este orden, y posible apomorfía (además de los datos moleculares), es la presencia de fitomelanina, un pigmento oscuro presente en las cubiertas seminales. Las fitomelaninas se encuentran en la mayoría de las familias de Asparagales, habiéndose perdido aparentemente en algunos taxones, siendo Orchidaceae el ejemplo más conspicuo en ese aspecto.
La mayoría de las especies de Asparagales son plantas herbáceas perennes, si bien hay algunos ejemplos de plantas trepadoras (ciertas especies de Asparagus, subfamilia Asparagoideae) y leñosas (algunos Aloe, subfamilia Asphodeloideae). Casi todas las especies de este orden producen un racimo apretado de hojas en la base de las plantas (son "plantas en roseta"), otras llevan esa misma disposición de hojas en el extremo de un tronco (Yucca), en algunos pocos casos las hojas se producen a lo largo del tallo. Las flores frecuentemente se hallan en la extremidad del tallo.
Dentro de las monocotiledóneas y desde el punto de vista económico, el orden Asparagales es el segundo en importancia después del orden Poales (el cual incluye a los pastos y los cereales). El orden contiene muchas especies de importancia hortícola (cebolla, ajo, espárrago, puerro), en floricultura (freesia, gladiolo, iris, muchos miembros de orquídeas), jardinería (Hemerocallis, Convallaria, Agapanthus) y especias (vainilla).
Desde el punto de vista de su taxonomía, Asparagales es un orden sugerido por Huber (1977), luego adoptado por Dahlgren et al. (1985), y reafianzado en 1998 por el Grupo de Filogenia de las Angiospermas (en inglés original, "Angiosperm Phylogeny Group", siglas APG), retomado en el 2003 por el APG II, y en el 2009 por el APG III), y también por el APWeb (2001 hasta ahora). Las delimitaciones de las familias de Asparagales han sufrido una serie de cambios en años recientes, y aún se necesitan más estudios para que se estabilice su circunscripción y sus relaciones.
Asparilaadsed (Asparagales) on õistaimede selts üheiduleheliste klassist.
Seltsi kuulub ühe süsteemi (nn. APG II süsteemi, 2003) järgi 16 sugukonda:
Cronquisti süsteemi järgi kuulub suur osa selle seltsi rühmi aga seltsi liilialaadsed (Liliales).
Asparilaadsed (Asparagales) on õistaimede selts üheiduleheliste klassist.
Seltsi kuulub ühe süsteemi (nn. APG II süsteemi, 2003) järgi 16 sugukonda:
Alliaceae Laugulised Asparagaceae Asparilised Asteliaceae Blandfordiaceae Boryaceae Doryanthaceae Hypoxidaceae Iridaceae Võhumõõgalised Ixioliriaceae Lanariaceae Orchidaceae Käpalised Tecophilaeaceae Xanthorrhoeaceae Asphodelaceae Aaloelised Hemerocallidaceae Päevaliilialised XeronemataceaeCronquisti süsteemi järgi kuulub suur osa selle seltsi rühmi aga seltsi liilialaadsed (Liliales).
Asparagales landare sailkapeneko taxoi bat da, gutxienez Asparagaceae familia barruan duena. Asparagales orain dela gutxi onartutako ordena bat da eta bere familia asko iraganean Liliales ordenean sartzen ziren. Liliales, hala ere, oso ordena zabala zen, ia monokotiledonea guztiak batzen zituena bere petaloak ikusgarriak baziren eta endosperman ez bazuen almidoirik. ADN analisietan oinarrituta ikusi zen Liliales ordenan Asparagales eta Dioscoreales gutxienez zeudela. Asparagalesen barruan 24 familia daude, eta 26.000 espezie inguru. Ordena ADN sekuentziazioan oinarritzne da, baina morfologiari dagokionez definitzea zaila da, estrukturalki oso anitza delako. Talde honen barruan 15 zentimetro baino ez duten belar batzuk sartzen dira eta Agave bezalakoak, 10 metroko altuera har dezaketenak.
Ordena definitzeko erabili daitekeen zerbait fitomelaninaren presentzia da, hazien oskolean agertzen den pigmentu ilun bat. Hala ere Orchidaceaen ez da agertzen pigmentu hau.
Asparagusen barruan badira landare igokariak direnak eta Aloe barruan zurezkoak direnak, baina gehienak belarkarak dira. Espezie gehienek hosto bilduma estu bat sortzen dute euren oinarrian. Normalki loreak zurtoinaren bukaeran aurki daitezke.
Monokotiledoneoen artean Poalesen ostean ekonomia garrantzi handiena duen taldea da. Tipula, baratxuria, zainzuria edo porrua talde honetan daude, baita ere orkidea asko zein banilla bezalako landareak.
Hemerocallidoideae (= Hemerocallidaceae)
Xanthorrhoeoideae (= Xanthorrhoeaceae s.s.)
Asphodeloideae (= Asphodelaceae)
Agapanthoideae (= Agapanthaceae)
Allioideae (= Alliaceae s.s.)
Amaryllidoideae (= Amaryllidaceae s.s.)
Aphyllanthoideae (= Aphyllanthaceae)
Brodiaeoideae (= Themidaceae)
Scilloideae (= Hyacinthaceae)
Agavoideae (= Agavaceae)
Lomandroideae (= Laxmanniaceae)
Asparagoideae (= Asparagaceae s.s.)
Nolinoideae (= Ruscaceae)
Asparagales landare sailkapeneko taxoi bat da, gutxienez Asparagaceae familia barruan duena. Asparagales orain dela gutxi onartutako ordena bat da eta bere familia asko iraganean Liliales ordenean sartzen ziren. Liliales, hala ere, oso ordena zabala zen, ia monokotiledonea guztiak batzen zituena bere petaloak ikusgarriak baziren eta endosperman ez bazuen almidoirik. ADN analisietan oinarrituta ikusi zen Liliales ordenan Asparagales eta Dioscoreales gutxienez zeudela. Asparagalesen barruan 24 familia daude, eta 26.000 espezie inguru. Ordena ADN sekuentziazioan oinarritzne da, baina morfologiari dagokionez definitzea zaila da, estrukturalki oso anitza delako. Talde honen barruan 15 zentimetro baino ez duten belar batzuk sartzen dira eta Agave bezalakoak, 10 metroko altuera har dezaketenak.
Ordena definitzeko erabili daitekeen zerbait fitomelaninaren presentzia da, hazien oskolean agertzen den pigmentu ilun bat. Hala ere Orchidaceaen ez da agertzen pigmentu hau.
Asparagusen barruan badira landare igokariak direnak eta Aloe barruan zurezkoak direnak, baina gehienak belarkarak dira. Espezie gehienek hosto bilduma estu bat sortzen dute euren oinarrian. Normalki loreak zurtoinaren bukaeran aurki daitezke.
Monokotiledoneoen artean Poalesen ostean ekonomia garrantzi handiena duen taldea da. Tipula, baratxuria, zainzuria edo porrua talde honetan daude, baita ere orkidea asko zein banilla bezalako landareak.
Asparagales on yksisirkkaisten (Liliopsida = ylälahko Lilianae) kasvien lahko, jonka heimojen on aiemmin katsottu kuuluvan lahkoon Liliales ja osan suvuista jopa heimoon Liliaceae. Jotkin järjestelmät erottavat ryhmästä omiksi lahkoikseen kämmekkämäiset kasvit (Orchidales) ja kurjenmiekkamaiset kasvit (Iridales). APG (Angiosperm Phylogeny Group) on kasvitieteilijöiden laajalti käyttämässä luokittelussaan sisällyttänyt lahkoon tässä artikkelissa mainitut heimot.
Asparagales-lahkon tyyppisuku on parsat (Asparagus). Sen ylivoimaisesti suurin heimo on kämmekkäkasvit (Orchidaceae), johon kuuluu noin 22 075 lajia. Muita merkittäviä heimoja ovat kurjenmiekkakasvit (2 025 lajia), parsakasvit (noin 2 480 lajia) ja narsissikasvit (1 605 lajia).[1]
Lahkoon kuuluu 14 heimoa, 1122 sukua ja 26070 lajia.[2]
Asparagales on yksisirkkaisten (Liliopsida = ylälahko Lilianae) kasvien lahko, jonka heimojen on aiemmin katsottu kuuluvan lahkoon Liliales ja osan suvuista jopa heimoon Liliaceae. Jotkin järjestelmät erottavat ryhmästä omiksi lahkoikseen kämmekkämäiset kasvit (Orchidales) ja kurjenmiekkamaiset kasvit (Iridales). APG (Angiosperm Phylogeny Group) on kasvitieteilijöiden laajalti käyttämässä luokittelussaan sisällyttänyt lahkoon tässä artikkelissa mainitut heimot.
Asparagales-lahkon tyyppisuku on parsat (Asparagus). Sen ylivoimaisesti suurin heimo on kämmekkäkasvit (Orchidaceae), johon kuuluu noin 22 075 lajia. Muita merkittäviä heimoja ovat kurjenmiekkakasvit (2 025 lajia), parsakasvit (noin 2 480 lajia) ja narsissikasvit (1 605 lajia).
Les Asparagales sont un ordre de plantes monocotylédones introduit par la classification phylogénétique des angiospermes. Il regroupe des familles dont beaucoup faisaient partie de l'ordre des Liliales ou résultent de l'éclatement de la famille des Liliacées.
En classification phylogénétique APG III (2009) :
En classification phylogénétique APG II (2003) :
N.B. « + ... » = famille optionnelle
La précédente liste est une modification apportée à la classification phylogénétique APG (1998) qui utilisait la liste suivante :
Ord de phlandaí is ea Asparagales, sna chórais aicmithe nua-aimseartha ar nós an Ghrúpa Fhíligineacht Angiospermae ( Béarla: Angiosperm Phylogeny Group) agus an Láithreáin Gréasáin Fhíligineacht Angiospermae (Béarla:Angiosperm Phylogeny Website).
Ord de phlandaí is ea Asparagales, sna chórais aicmithe nua-aimseartha ar nós an Ghrúpa Fhíligineacht Angiospermae ( Béarla: Angiosperm Phylogeny Group) agus an Láithreáin Gréasáin Fhíligineacht Angiospermae (Béarla:Angiosperm Phylogeny Website).
.Asparagales é unha orde de plantas anxiospermas utilizada en sistemas de clasificación modernos como o sistema de clasificación APG III do ano 2009[3] e o APWeb,[4] que comprende polo menos a familia Asparagaceae. Asparagales é unha orden que se recoñeceu moi recentemente nos sistemas de clasificación e moitas das súas familias pertencían á antiga orde Liliales. Liliales fora unha orde moi ampla na cal se situaban todas as monocotiledóneas de tépalos vistosos e sen amidón no endosperma, que fora difícil de dividir en familias porque os seus caracteres morfolóxicos non aparecían en padróns que delimitasen claramente grupos. Co tempo empezouse a recoñecer a orde Asparagales, especialmente cando as análises moleculares de ADN indicaron que Liliales debía ser dividida polo menos en Liliales, Asparagales e Dioscoreales. Asparagales é unha orde bastante grande, consta de 14 familias e unhas 26 000 especies. Aínda que a orde está claramente circunscrita baseándose nas análises de secuencias de ADN pero é dificultosa de definir morfoloxicamente, xa que é un dos grupos de monocotiledóneas estruturalmente máis diverso. Así, a orde inclúe desde plantas herbáceas de non máis de 15 cm ata algunhas especies de Agave que superan os 10 m de altura. Nestes sentido, a súa separación da orde Liliales é particularmente problemática.
Unha das características que definen esta orde, e posible apomorfía (ademais dos datos moleculares), é a presenza de fitomelanina, un pigmento escuro presente nas cubertas das sementes. As fitomelaninas encóntranse na maioría das familias de Asparagales, pero perdéronse aparentemente nalgúns taxons, e o exemplo máis salientable nese aspecto é Orchidaceae.
A maioría das especies de Asparagales son plantas herbáceas perennes, aínda que hai algúns exemplos de plantas gabeadoras (certas especies de Asparagus, subfamilia Asparagoideae) e leñosas (algúns Aloe, subfamilia Asphodeloideae). Case todas as especies desta orde producen unha roseta apertada de follas na base das plantas (son "plantas en roseta"), outras levan esa mesma disposición de follas no extremo dun tronco (Yucca), e nuns poucos casos as follas aparecen ao longo do talo. As flores frecuentemente están no extremo do talo.
Dentro das monocotiledóneas e desde o punto de vista económico, a orde Asparagales é a segunda en importancia despois da orde Poales (a cal comprende os pastos e os cereais). A orde contén moitas especies de importancia hortícola (cebola, allo, espárrago, porro), en floricultura (Freesia, gladíolo, iris, moitas orquídeas), xardinería (Hemerocallis, Convallaria, Agapanthus) e especias (vainilla).
Desde o punto de vista da súa taxonomía, Asparagales é unha orde suxerida por Huber (1977),[5] logo adoptado por Dahlgren et al. (1985),[6] e reafianzada en 1998 polo Grupo de Filoxenia das Anxiospermas (en inglés orixinal, "Angiosperm Phylogeny Group", siglas APG),[7] retomado no 2003 polo APG II,[8] e en 2009 polo APG III),[1] e tamén polo APWeb (2001 ata agora).[4] Porén, as delimitacións das familias de Asparagales sufriron unha serie de mudanzas en anos recentes, e aínda se necesitan máis estudos para que se estabilice a súa circunscrición e as súas relacións.
A maioría das especies de Asparagales son plantas herbáceas perennes; hai tamén algúns exemplos de plantas gabeadoras (certas especies de Asparagus, subfamilia Asparagoideae das asparagáceas) e leñosas (algúns Aloe, subfamilia Asphodeloideae das xantorroeáceas). Case todas as especies desta orde producen un conxunto de follas na base das plantas (son "plantas en roseta"), outras presentan esa mesma disposición de follas no extremo dun tronco (Yucca), e nuns poucos casos as follas están dispostas ao longo do talo. As flores frecuentemente están no extremo do talo. Asparagales pode ser distinguido de Liliales polos seus tépalos xeralmente non punteados, nectarios nos septos dos ovarios (en lugar de na base dos tépalos ou estames) e ás veces un crecemento secundario anómalo (a diferenza da falta de crecemento secundario). Os membros de Asparagales adoitan ser xeófitas, pero neste caso as súas follas son en xeral lineais e sen venación reticulada fina.
As súas sementes son características, coa epiderme externa da cuberta seminal obliterada (na maioría das especies de froitos carnosos), ou presente e cunha costra de fitomelaninas negra e carbonácea en moitas especies de froito seco. Normalmente, a parte interna da cuberta seminal está completamente colapsada. En contraste, as sementes de Liliales (morfoloxicamente similares) sempre teñen unha epiderme externa ben desenvolvida, non teñen fitomelaninas, e usualmente reteñen unha estrutura celular na porción interna da cuberta seminal.
O carácter infrecuente de ter un crecemento secundario anómalo cun meristema en ringleiras superpostas ("etáxeno") está prácticamente restrinxido aos xéneros deste clado (fóra deste clado, está presente só en Dioscorea). Algunhas destas especies, dos xéneros Agave, Aloe, Cordyline, Dracaena, Nolina e Yucca, convértense en árbores curtas e macizas, xeralmente (pero non sempre) con ramificación limitada (por exemplo Aloe, Dracaena, Yucca). Outros xéneros como Aphyllanthes e Lomandra, teñen este mesmo tipo de crecemento secundario pero restrinxido aos seus talos subterráneos.
Outros caracteres apomórficos da orde son: a presenza de ácido chelidónico, quizais a de velame, talvez a das células hipodérmicas das raíces dimórficas, as anteras máis longas que largas, as células do tapete de bi- a tetranucelares, a microsporoxénese simultánea (pero ver o apartado de Filoxenia), a presenza de estilo no pistilo, as sementes con exotesta, o tegmen non persistente, o endosperma helobial, e a perda do xene mitocondrial sdh3.
Se a microsporoxénese simultánea é unha apomorfía da orde, no "Núcleo dos Asparagales" aparentemente cambiou a sucesiva. Hai taxons de clados máis basais onde a microsporoxénese é sucesiva, nos que os detalles da formación da parede son similares aos dos membros do núcleo dos Asparagales que foron estudados: placas celulares centrífugas. Pero nos membros basais nos que a microsporoxénese é simultánea, a formación das láminas tamén pode ser centrifuga (Nadot et al. 2006).[9]
Algúns autores consideran o ovario ínfero unha sinapomorfía da orde, pero en caso de que o fose, o carácter perdeuse tantas veces que resulta ser un carácter moi lábil (dentro da orde coma no resto das anxiospermas), polo que a posición do ovario dentro da orde non ten valor preditivo.
Encóntranse tépalos de 3 trazas en Orchidaceae, Amaryllidoideae, Iridaceae, Asphodeloideae (mais non en Kniphofia, Asphodelus), Agavoideae, Agapanthoideae e Hemerocallidoideae; encóntranse tépalos dunha traza en Nolinoideae (mais non en Maianthemum stellatum), Allioideae, Aphyllanthoideae e Asparagoideae. Scilloideae ten tépalos cos dous tipos de vasculatura, incluso se encontran tépalos de 5 trazas nos verticilos externos de Urginea.
Fixéronse estudos dos telómeros das especies da orde. Parece que Asparagales representa unha situación dinámica na cal se mantén unha variedade de motivos teloméricos en baixa cantidade de copias, en lugar da repetición dominante de tipo Arabidopsis. Parece que nalgúns casos, un dos tipos que estaba en baixa cantidade de copias é amplificado e substitúe o tipo estándar. Por exemplo, sabíase desde hai moitos anos que Allium non posuía as repeticiones teloméricas estandarizadas de tipo Arabidopsis e logo descubriuse[10] que Aloe tampouco as tiña, polo que se predixo que os xéneros intermedios tampouco as terían: nunha análise feita a representantes de todos os Asparagales descubriuse que, agás Ornithogalum, ningún tiña as repeticións tipo Arabidopsis. Máis adiante[11] descubriuse que Ornithogalum e outros xéneros substituíran as repeticións estándar das plantas por unhas de tipo humano, e que practicamente todos os xéneros de Asparagales teñen polo menos algunhas copias das repeticións teloméricas de tipo humano, como tamén unha variedade doutras repeticións situadas nas terminacións dos seus cromosomas. A presenza e a composición desta variedade de repeticións teloméricas posúen un gran valor para discernir a árbore filoxenética.
Na orde encontráronse xenomas moi grandes, cun valor C duns 350 picogramos, nalgunhas Allioideae, Amaryllidoideae, Scilloideae e Orchidaceae.[12]
En resumo, a anatomía da semente parece posuír o mellor conxunto de caracteres que unifican a orde, que noutros aspectos é moi heteroxéneo (particularmente tendo en conta que Orchidaceae está dentro del).
As Asparagales comunmente presentan micorrizas arbusculares de tipo Arum, mentres que en Liliales estas micorrizas son normalmente de tipo Paris.[13]
A diversidade taxonómica das monocotiledóneas foi presentada en detalle por Kubitzki (1998,[14] 2006).[15]
Velaquí unha lista da diversidade de Asparagales, onde se mostran diversas familias e subfamilias. As descricións son breves e para máis información hai que consultar as ligazóns.
A monofilia de Asparagales está apoiada en análises cladísticas baseadas en morfoloxía, o ADNr de 18S e moitas secuencias de ADN (Chase et al. 1995a,[20] b,[21] 2000,[22] 2006,[23] Conran 1989,[24] Davis et al. 2004,[25] Graham et al. 2006,[26] Hilu et al. 2003,[27] Pires et al. 2006,[28] Rudall et al. 1997,[29] Källersjö et al. 1998,[30] Fay et al. 2000,[31] Soltis et al. 2000,[32] Stevenson et al. 2000,[33] McPherson e Graham 2001),[34] aínda que algunhas reconstrucións filoxenéticas como a de Hilu et al. (2003),[27] con datos moleculares, suxeriron que Asparagales pode ser parafilético, coas Orchidaceae separadas do resto.
Con respecto ás relacións de Asparagales co resto das ordes de monocotiledóneas, é o clado irmán do clado Commelinidae.
Segundo as análises morfolóxicas feitas por et al. (1995b)[21] e Stevenson e Loconte (1995),[35] polo menos algunhas das familias de Asparagales con crecemento secundario anómalo formaban clados, aínda que tamén quedaban dentro destes clados algúns membros que hoxe están situados en Liliales, Zingiberales e Pandanales. Outro traballo recente de filoxenia da orde con datos morfolóxicos é o de Rudall (2002).[36]
As relacións dentro da orde foron investigadas por Dahlgren et al. (1985),[6] Rudall e Cutler (1995),[37] Chase et al. (1995a,[20] b,[21] 1996),[38] Stevenson e Loconte (1995),[35] Rudall et al. (1997),[29] Fay et al. (2000),[31] McPherson e Graham (2001),[34] Graham et al. (2006)[26] e Pires et al. (2006).[28]
Velaquí o cladograma resumido, segundo Judd et al. 2007 (modificado de Soltis et al. 2005,[39] con Orchidaceae basal como en APWeb 2008):[4]
AsparagalesHypoxidaceae (tradicionalmente situado en Amaryllidaceae)
Xanthorrhoeaceae sensu latoHemerocallidoideae (tradicionalmente situado en Liliaceae)
Xanthorrhoeoideae (tradicionalmente situado en Liliaceae)
Asphodeloideae (tradicionalmente situado en Liliaceae)
Asparagales superiores Amaryllidaceae sensu latoAgapanthoideae (tradicionalmente situado en Liliaceae)
Allioideae (tradicionalmente situado en Liliaceae)
Lomandroideae (antes familia Laxmanniaceae, tradicionalmente situado en Liliaceae)
Asparagoideae (tradicionalmente situado en Liliaceae)
Nolinolidease (antes familia Ruscaceae, tradicionalmente situado en Liliaceae)
Brodiaeoideae (antes familia Themidaceae, tradicionalmente situado en Liliaceae)
Scilloideae (antes Hyacinthaceae, tradicionalmente situado en Liliaceae)
Agavoideae (tradicionalmente situado en Amaryllidaceae ou Liliaceae)
A APWeb proporciona este cladograma completo:[4]
Asparagales Hypoxidaceae sensu lato? Xanthorrhoeaceae sensu lato Asparagales superiores (= Núcleo dos Asparagales) Amaryllidaceae sensu lato Asparagaceae sensu latoBrodiaeoideae = Themidaceae
Scilloideae = Hyacinthaceae
Agavoideae (Anemarrhena, Agave e afíns, Hesperocallis, Behnia, Herreria e afíns, Anthericum e afíns)
Lomandroideae = Laxmanniaceae
Nolinoideae = Ruscaceae
A árbore, cos nomes dos clados actualizados segundo o APG III (2009),[1] está baseada principalmente nas análises de Chase et al. (2000)[22] e Fay et al. (2000).[31] Estes estudos difiren pouco nos detalles, pero a análise de Fay et al. (2000)[31] ten máis nodos con bo apoio no núcleo dos Asparagales (o cal non é sorprendente, xa que utilizaron o método de ponderación sucesiva). Tamén é moi congruente con esta filoxenia a presentada por McPherson e Graham (2001),[34] feita cunha mostraxe moito menor.
Das análisis despréndese unha ensamblaxe basal parafilética, chamada doe "Asparagales inferiores" (Lower Asparagales sensu Rudall et al. 1997),[29] que inclúe a familias como Orchidaceae, Hypoxidaceae, Iridaceae, Asphodeloideae e Hemerocallidoideae. En contraste, os chamados "Asparagales superiores" (Higher Asparagales) son un clado propiamente dito que se separa do resto, e inclúe a Allioideae, Amaryllidoideae, Scillloideae = Hyacinthaceae, Agavoideae, Asparagoideae e Nolinoideae = Ruscaceae. Moitos dos "Asparagales inferiores" caracterízanse pola microsporoxénese simultánea (as 4 micrósporas sepáranse á vez despois das dúas divisións meióticas). O padrón de microsporoxénese dos "Asparalales inferiores" é apomórfico dentro das monocotiledóneas. O padrón de microsporoxénese dos "Asparagales superiores", ao contrario, é como os das monocotiledóneas máis basais. Rudall (2001,[40] ver tamén 2002b,[41] 2003a)[42] incluíu o carácter do ovario ínfero como unha sinapomorfía da orde, notando que nos "Asparagales superiores" puido terse producido unha reversión ao ovario súpero, reversión asociada coa presenza de nectarios septais infraloculares (como en Xanthorrhoea e Johnsonia, dous xéneros de Hemerocallidoideae). Porén, Stevens (2001 en adiante, APWeb)[4] nota que os ovarios súperos tamén aparecen distribuídos nos "Asparagales inferiores", polo que non está claro onde deben localizarse na árbore as evolucións das diferentes morfoloxías do ovario: a posición do ovario parece un carácter moito máis flexible (aquí coma nas demais anxiospermas) do que se cría. Os "Asparagales superiores", por seren un grupo monofilético con forte apoio nas análises moleculares de ADN, denomínanse "Núcleo dos Asparagales" ("Core Asparagales") no APWeb.[4]
A familia Orchidaceae é a máis grande de todas as anxiospermas. Algúns autores (por exemplo Dahlgren et al. 1985)[6] recoñeceron tres familias de orquídeas, pero as análises moleculares de ADN demostraron que estas familias son polifiléticas e deberían reunirse nun taxon Orchidaceae sensu lato, que é como se trata a familia hoxe en día. Con respecto á súa relación coas demais familias de Asparagales, varios traballos recentes suxiren que Orchidaceae é irmán do resto dos Asparagales (por exemplo con apoio bootstrap do 76 % en Graham et al. 2006,[26] un resultado máis ou menos similar en Givnish et al. 2006b,[43] un apoio máis forte -do 96 ao 99 %-, cun bo tamaño de mostraxe e 7 xenes estudados, en Pires et al. 2006,[28] e tamén un bo apoio en Chase et al. 2005,[44] onde o nodo que separa as orquídeas do resto de Asparagales tiña un 91 % de apoio bootstrap). Outros traballos situárona de forma diferente na árbore filoxenética: ás veces situada como irmán de Boryaceae (por exemplo en Chase et al. 1995a[20] e en McPherson e Graham 2001,[34] pero cun apoio máis ben baixo). En cambio Li e Zhou (2007)[45] consideran Orchidaceae embebida nun clado parafilético Boryaceae-Hypoxidaceae. Por outro lado, Rudall (2003a),[42] con datos morfolóxicos, suxire unha relación morfolóxica próxima entre Hypoxidaceae e Orchidaceae en particular. Porén, Janssen e Bremer (2004),[46] non sitúan Orchidales como irmán do resto da orde, senón temporalmente preto do comezo da diverxencia dentro dela (aínda que a topoloxía da árbore que utilizaron difire considerablemente en detalle da utilizada aquí). En resumo, a posición de Orchidaceae como irmán do resto de Asparagales, con Boryaceae como parte do clado que inclúe Blandfordiaceae, parece a mellor hipótese, e esta topoloxía é a presentada aquí. Nótese que isto cambia a caracterización de Asparagales; moitos dos seus caracteres agora foron resituados no nodo subbasal no clado (comparar a lista de apomorfías do APWeb[4] nas versións 7 e anteriores a esa do sitio web). Un dato interesante é que as orquídeas carecen da maioría das sinapomorfías non moleculares da orde: as sementes de Orchidaceae non teñen fitomelaninas debido á súa natureza (son pulverulentas), e os seus nectarios raramente están nos septos dos ovarios. As orquídeas teñen microsporoxénese simultánea e ovario ínfero, caracteres que son típicos dos "Asparagales inferiores" de Rudall et al. 1997,[29] mais non está claro se estes caracteres deberían considerarse sinapomorfías da orde. Aparentemente a única sinapomorfía de Orchidaceae é a presenza de protocormo (a estrutura producida polo crecemento dos seus embrións indiferenciados antes de que se desenvolvan as raíces e talos). As orquídeas presentan o androceo e o xineceo fusionados, pero esta característica está presente só na subfamilia Apostasioideae e só en graos variables. As sementes similares a po dispersadas polo vento están presentes na maioría das subfamilias, pero nalgúns membros de Cypripedioideae e Vanilloideae hai sementes crustosas (probablemente asociadas á súa forma de dispersión, que neste caso é relizada por paxaros e mamíferos, que son atraídos polo seu froito carnoso que fermenta in situ liberando compostos fragantes, por exemplo a vainillina). A diversificación en Orchidaceae é remarcable, mais debe compararse coa dos demais Asparagales, que son menos ricos en especies pero moi variables morfoloxicamente.
As relacións de Boryaceae (que comprende só dous xéneros, Borya e Alania) co resto das familias de Asparagales permaneceron pouco claras durante moito tempo. As boryáceas son micorrícicas, pero non como as orquídeas, senón que as súas micorrizas son do tipo estandarizado vesicular-arbuscular (VA). Previamente chegárase a pensar que Boryaceae era un membro de Anthericaceae (Dahlgren et al. 1985,[6] Takhtajan 1997),[47] unha familia que demostrou ser extremadamente polifilética (Chase et al. 1996).[38] A situación de Orchidaceae como irmán de todo o resto de Asparagales dálle un apoio relativaemnte importante á situación de Boryaceae como grupo irmán do clado que inclúe a Blandfordiaceae. Rudall (2003a)[42] tamén suxeriu unha relación morfolóxica próxima entre Boryaceae e Blandfordiaceae. Porén, hai que ter en conta que aínda que hai un bo apoio na árbore de Chase et al. (2006)[23] para posicionar Orchidaceae como irmán de todas as demais Asparagales, Boryaceae está situado inmediatamente por encima do clado que inclúe Blandfordiaceae, pero cun apoio baixo.
Hypoxidaceae é unha familia que tradicionalmente fora colocada en Amaryllidoideae. Por outr parte, Lanariaceae co seu único xénero Lanaria foi no pasado considerado un membro de Haemodoraceae (Hutchinson 1967)[48] ou Tecophilaeaceae (Dahlgren et al. 1985).[6] Estas dúas familias xunto con Asteliaceae, Blandfordiaceae e probablemente tamén Boryaceae forman un clado, e excepto pola situación de Boryaceae, as outras 4 teñen bo apoio como clado nas análises moleculares. Unha sinapomorfía potencial para estas familias é a estrutura do óvulo: polo menos Asteliaceae, Blandfordiaceae, Lanariaceae e Hypoxidaceae posúen unha constrición calazal e unha cofia nucelar. Soltis et al. (2005), deixando fóra do clado a Boryaceae, suxeriron que estas familias se fusionaran nun taxon Hypoxidaceae sensu lato.
Dentro deste clado, Asteliaceae + Hypoxidaceae + Lanariaceae forman á súa vez un clado con apoio do 100 % de bootstrap. A morfoloxía tamén proporciona algún apoio para estas relacións: Asteliaceae e Hypoxidaceae forman rosetas cubertas con pelos multicelulares ramificados (tamén Lanariaceae posúe pelos multicelulares), e posúen canais radiculares cheos de mucílago (este último carácter está tamén presente en Lanariaceae), pero Blandfordia (único xénero de Blandfordiaceae) non comparte estes caracteres. Ademais, este clado posúe estomas paracíticos, a lámina da folla cunha vea media distintiva, quizais o xineceo máis ou menos ínfero, e un micrópilo biestomal. Asteliaceae + Hypoxidaceae posúen flavonois, endosperma de paredes delgadas, cotiledóns non fotosintéticos, e a lígula longa.
O seguinte nodo en Asparagales (todas as familias menos Orchidaceae e o clado mencionado) demostrouse que é robusto (81 % de apoio jacknife en Soltis et al. 2000,[32] 100 % de apoio bootstrap en Chase et al. 2005,[44] ver tamén Fay et al. 2000[31] usando datos de catro xenes plastídicos, e Chase et al. 2000[22]), pero non hai caracteres fóra dos moleculares que unan a este clado.
A posición de Ixioliriaceae e Tecophilaeaceae aínda é pouco clara. Algunhas análises moleculares únenas nun clado, por exemplo recibe apoio de leve a moderado en Chase et al. (2000),[22] Pires et al. (2006),[28] e Givnish et al. (2006),[43] e en Graham et al. (2006)[26] recibe un apoio máis forte pero a mostraxe é máis ben pobre. Na análise de 3 xenes feita por Fay et al. (2000)[31] o clado ten un brazo moi longo. Estas dúas familias comparten a posesión de cormos, unha inflorescencia foliosa e moitas veces case capitada, follas de disposición espiralada e base envaiñadora, flores máis ben grandes, verticilo externo de tépalos mucronado a aristado, perianto formando un tubo curto, androceo inserido na boca, e un número cromosómico x=12 (Soltis et al. 2005 e APWeb). Tamén se encontran outras topoloxías: en Janssen e Bremer (2004)[46] Ixioliriaceae diverxe considerablemente antes (pero é adxacente a) Tecophilaeaceae; e en Davis et al. (2004),[25] cunha mostraxe menor, encóntrase algún apoio para irmandar a Ixioliriaceae con Iridaceae, tamén en Chase et al. (2006)[23] hai apoios (esta vez fortes) para este parentesco, onde o clado é irmán de Doryanthaceae (mais con apoio baixo).
As relacións de Doryanthaceae coas familias que a rodean nas análises filoxenéticas aínda son pouco claras tanto nas análises morfolóxicas coma nas moleculares. Chase et al. (2006)[23] colocárona na posición que se acaba de mencionar, que non é a presentada aquí. Para a situación presentada aquí hai apoio moderado en Fay et al. (2000),[31] e un apoyo do 92 % en Graham et al. (2006).[26] Rudall (2003)[42] que atopou unha relación morfolóxica próxima entre Doryanthaceae e Iridaceae.
O nodo que contén Iridaceae e todo o resto de Asparagales (Xeronemataceae etc.) recibe un apoio bastante forte en Fay et al. (2000).[31] O grupo ten algunhas características moleculares en común: perdeu os telómeros de tipo Arabidopsis, mentres que son comúns os telómeros de tipo humano (TTAGGG).
Iridaceae é unha das familias máis grandes e mellor estudadas de Asparagales (por exemplo ver Goldblatt 1990,[49] 1991,[50] Goldblatt et al. 1998,[51] 2002,[52] Reeves et al. 2001).[53] Iridaceae é distintiva entre os Asparagales debido á estrutura única da súa inflorescencia (un ripidio) e a súa combinación de ovario ínfero con 3 estames; tamén son comúns na familia as follas unifaciais, cando as follas bifaciais son a norma noutros Asparagales.
O nodo que segue á posible separación de Iridaceae está ben sostido nas análises (Fay et al. 2000,[31] Chase et al. 2005).[44] Se ter un ovario ínfero é una sinapomorfía de Asparagales (Chase et al. 1995b),[21] entón este nodo marcaría a transición a un ovario súpero (aínda que se encontran ovarios ínferos en Amaryllidoideae e en Yucca). Todos os clados deste nodo teñen nectarios septais infraloculares, o que Rudall (2000)[54] interpreta como ovarios secundariamente súperos. Esta interpretación demostra a reversibilidade deste carácter que moitas veces foi enfatizado na sistemática de monocotiledóneas (por exemplo en Cronquist 1981,[55] Dahlgren et al. 1985).[6] É neste nodo no que se perde o xene mitocondrial rpl2 (ou quizais no seguinte, de acordo coa distribución de Adams et al. 2002).[56]
Xeronemataceae co seu único xénero Xeronema foi previamente considerado como estreitamente emparentado con Phormium (usualmente Phormiaceae, Dahlgren et al. 1985),[6] pero aínda que hai similitudes superficiais (como as follas unifaciais), non son idénticos nos detalles. Chase et al. (2000)[22] describiu recentemente Xeronemataceae, e os seus caracteres varían segundo padróns que son xerais en Asparagales.
O seguinte nodo da árbore (Xanthorrhoeaceae sensu lato + os "Asparagales superiores") ten forte apoio en Fay et al. (2000)[31] e en Chase et al. (2000c).[57]
Hemerocallidoideae está próxima a Asphodelaceoideae e Xanthorrhoeoideae, e os 3 clados teñen antraquinonas, estiloides e sementes con cotiledón non fotosintético. Ver Kite et al. (2000)[58] para a distribución de antraquinonas, e McPherson et al. (2004)[59] para os taxons que non teñen o intrón 3'-rps12. Dentro deste clado, dáse crecemento secundario anómalo en Aloe (algunhas especies chegan a tamaños enormes), Phormium e Xanthorrhoea. As 3 familias son reunidas por algúns autores nun taxon Xanthorrhoeaceae sensu lato (como en APG III e no APWeb, 2001 en adiante),[4] aínda que Stevens no APWeb remarca que a microsporoxénese é variable neste grupo. En Fay et al. (2000)[31] encóntrase forte apoio para este clado. Porén, as relacións de parentesco dentro del aínda non están esclarecidas. Na análise de 3 xenes de Chase et al. (2000)[22] hai un apoio moderado para a relación (Xanthorrhoeoideae + Asphodeloideae), que entre os caracteres morfolóxicos encontra a presenza de engrosamento secundario, o androceo non adnato aos tépalos, a presenza de hipostase, e as sementes angulosas. Esta relación é a presentada aquí, porén, o apoio diminúe cando se inclúen os taxons onde faltan algunhas secuencias de xenes, ver tamén Fay et al. 2000).[31] Porén, Devey et al. (2006)[60] encontran algún apoio para xuntar Xanthorrhoeoideae con Hemerocallidoideae (ver tamén Pires et al. 2006),[28] mentres que Chase et al. (2006)[23] suxiren un clado formado por Asphodeloideae + Hemerocallidoideae (pero a mostraxe é cuestionada no APWeb).[4] Con respecto á morfoloxía, tanto Hemerocallidoideae coma Xanthorrhoeoideae teñen ovarios que probablemente son secundariamente súperos e teñen nectarios septais infraloculares (Rudall 2002,[36] 2003).[42] Rudall (2003)[42] suxire unha relación morfolóxica próxima entre Hemerocallidoideae e Asphodeloideae, e outra entre Xanthorrhoeoideae e Iridaceae.
O valor das antraquinonas como carácter taxonómico é aprezado agora, xa que Asphodeloideae as posúe pero anteriormente non fora situada como próxima ás outras dúas familias de Xanthorrhoeaceae sensu lato senón a Anthericaceae (coa cal agora se sabe que está distantemente relacionada). Debido á presenza de antraquinonas Asphodeloideae foi situada separada de Anthericaceae por Dahlgren et al. (1985),[6] ademais de que Asphodeloideae ten microsporoxénese simultánea mentres que Anthericaceae ten microsporoxénese sucesiva. Asphodeloideae ten moitos xéneros con cariotipos bimodais distintivos (Chase et al. 2000c),[57] e este carácter utilizouse como base para suxerir que estes xéneros se separasen nunha familia Aloaceae (Dahlgren et al. 1985).[6] Porén, os estudos moleculares mantiveron que Asphodeloideae quedara na súa circunscrición ampla.
Xanthorrhoeoideae antes incluía outros taxons arborescentes como Kingia (que non ten crecemento secundario e agora é situado en Dasypogonaceae) e Cordyline (co cal está só distantemente emparentado, e é un membro de Lomandroideae, antes Laxmanniaceae).
Hemerocallidaceae moitas veces comprendía só o xénero Hemerocallis (Dahlgren et al. 1985),[6] pero xa desde a circunscrición do APG (1998),[7] a familia incluía un gran número de xéneros, como Dianella, Johnsonia, e Phormium. Todos estes xéneros teñen pole tricotomosulcado, unha rareza entre as monocotiledóneas, e os estudos moleculares temperáns (Chase et al. 1995a)[20] foron críticos en determinar se este carácter era importante para estes xéneros (Rudall et al. 1997).[29] No APG III é a subfamilia Hemerocallidoideae de Xanthorrhoeaceae sensu lato.
O resto das familias, que se poderían reunir nos clados Amaryllidaceae sensu lato e Asparagaceae sensu lato, denomínase en conxunto "Núcleo dos Asparagales" polo APWeb,[4] caracterízase por posuír microsporoxénese sucesiva (son os "Asparagoides superiores" de Rudall et al. 1997),[29] e quizais tamén por posuír saponinas esteroides. Nas análises moleculares de ADN é un grupo con forte apoio (por exemplo en Chase et al. 1995a,[20] Fay et al. 2000,[31] Chase et al. 2000c,[57] Graham et al. 2005).[61] Porén, a inclusión de Aphyllanthoideae (en Asparagaceae sensu lato), tende a diminuír o apoio (Graham et al. 2006).[26] Ademais, hai pouco apoio morfolóxico especialmente para as agrupacións dentro da área de Asparagaceae.
Amaryllidaceae sensu lato (Allioideae, Amaryllidoideae e Agapanthoideae) ten en común as semellanzas nas súas secuencias de ADNcp (Fay et al. 2000,[31] pero ver McPherson et al. 2004[59] e Thomas et al. 2005) e as súas secuencias de ADNnr ITS para Amaryllidoideae + Agapanthoideae (Meerow et al. 2000.[62]
Dentro do clado, a agrupación [Agapanthoideae [Allioideae + Amaryllidoideae]] foi suxerida por Meerow et al. (1999),[63] Fay et al. (2000,[31] con forte apoio), Givnish et al. (2006)[43] e Pires et al. (2006),[28] pero Meerow et al. (2000a)[64] consideraron Agapanthoideae como irmán de Amaryllidoideae, pero con baixo apoio.
O clado posúe algúns caracteres morfolóxicos: as inflorescencias son escaposas, umbeloides (construción cimosa), xeralmente con dúas (ou máis) brácteas como espatas escariosas fusionadas que as encerran. As follas son dísticas, os pedicelos non articulados, o estilo longo, o estigma seco, o endospermo nuclear ou helobial, sen hipocótilo, quizais deberían incluírse os tépalos libres, as anteras conadas na base e as células do tapete uninucleadas. (Allioideae + Amaryllidoideae) posúe como posibles sinapomorfías o feito de que son xeófitas, o talo que é un bulbo de ramificación simpodial, tunicado, e as raíces contráctiles.
As 7 familias que forman o clado Asparagaceae sensu lato están ben sostidas na análise de Fay et al. (2000,[31] sen incluír Hesperocallis), pero non hai caracteres morfolóxicos moi evidentes comúns ao clado, aínda que é bo candidato o endosperma: helobial, de paredes grosas, hemicelulósico. Incluso as familias en sentido estrito deste clado son difíciles de recoñecer, porque non posúen sinapomorfías obvias e as súas flores teñen o típico aspecto das liliáceas (nese sentido o APWeb[4] suxire que podería ser útil reunilas nun grupo Asparagaceae sensu lato, como de feito fai o APG III en 2009). Para detalles das súas relacións, ver Jang e Pfosser (2002)[65] e Bogler et al. (2006).[66]
En Asparagaceae sensu lato, a diferenza de Amaryllidaceae sensu lato, as inflorescencias son acios (pero Brodiaeoideae = Themidaceae ten inflorescencias umbeloides pero neste caso, teñen brácteas por debaixo de todas as flores, polo que as flores se interpretan como acios condensados). En Asparagaceae sensu lato hai moitos xéneros arborescentes (como Yucca, Agave, Cordyline), e tamén algúns con raíces con crecemento secundario (como Herreria, Thysanotus), e moitos son xeófitas. Amaryllidaceae sensu lato é moito menos diversa, e a maioría son xeófitas, bulbosas, pero algunhas son rizomatosas (como Agapanthus, Tulbaghia, Clivia). Un carácter morfolóxico que une as asparagáceas sensu lato é a presenza dunha inflorescencia racimosa.
Brodiaeoideae = Themidaceae, Scilloideae = Hyacinthaceae, Aphyllanthoideae e Agavoideae forman un clado (ver Fay et al. 2000[31] para todas menos Aphyllanthoideae), pero o sostén morfolóxico para este grupo é pouco claro. Este clado pode ser irmán do clado (Nolinoideae = Ruscaceae + Asparagoideae + Lomandroideae = Laxmanniaceae).
As relacións entre Aphyllanthoideae, Brodiaeoideae = Themidaceae e Scilloideae = Hyacinthaceae probablemente deberían representarse como unha tricotomía. Fay et al. (2000),[31] Pires et al. (2001),[67] e Pires e Sytsma (2002)[68] discuten a incerteza con respecto ás relacións de Brodiaeoideae. (Brodiaeoideae + Scilloideae), que parecen ter un moderado sostén en Fay e Chase (1996)[69] e en Meerow et al. (2000),[64] pero o apoio é baixo na análise de 2 xenes de Jang e Pfosser (2002,[65] sen incluír Aphyllanthes), en Chase et al. (2006),[23] e en Pires et al. (2006).[28] Os dous clados comparten as saponinas esteroides, as follas de disposición espiralada, os pedicelos bracteados, o óvulo anátropo, o endosperma helobial ou nuclear, e o cotiledón non fotosintético. Para algúns caracteres máis ver Fay e Chase (1996).[69] Nos dous clados poden estar presentes estruturas parecidas a laticíferos.
Aphyllanthes ten unha rama moi longa na análise de 3 xenes de Fay et al. (2000),[31] e a súa posición filoxenética é pouco clara, pero a súa eliminación dalgunhas análises diminúe drasticamente o apoio (Chase et al. 2006).[23] McPherson e Graham (2001) encontraron unha relación de Aphyllantes como irmán de Scilloideae,[34] pero Pires et al. (2006)[28] considerárono como irmán de Lomandroideae, pero con apoio baixo.
As subfamilias Lomandroideae + Asparagoideae + Nolinoideae comparten as saponinas esteroides, os pedicelos articulados, o froito tipo cápsula, o endosperma helobial, de paredes grosas, con puntuacións, hemicelulósico. Hai apoio moderado para este clado na árbore de 4 xenes do cloroplasto de Fay et al. (2000).[31]
Finalmente, Nolinoideae e Asparagoideae poden estar relacionadas, como se evidencia polos seus froitos indehiscentes, as súas flores bastante pequenas e o seu número cromosómico x=10; esta relación recibiu apoio prelilminar nalgunhas análises de secuencias de ADN (Chase et al. 1995a,[20] b,[21] 1996,[38] 2000,[22] Fay et al. 2000[31]). Os froitos carnosos con sementes sen fitomelaninas son comúns neste clado, pero non se sabe se este carácter constitúe unha apomorfía. Posúen froitos capsulares Hemiphylacus (irmán do resto de Asparagoideae) e o clado (Comosperma + Eriospermum), que é irmán do resto de Nolinoideae, polo que pode ser que o froito carnoso sexa un carácter derivado dentro do clado.
Para o desenvolvemento dos filóclados en Asparagus, Ruscus, e afíns, ver Cooney-Sovetts e Sattler (1987).[70]
O sistema APG de 1998[7] e o APG II de 2003[8] recoñeceron a Hesperocallidaceae co seu único xénero Hesperocallis, pero foi situado en Agavaceae polas últimas análises moleculares de ADN (Pires et al. 2004),[71] por iso a familia foi incluída en Agavaceae no APG III de 2009[1] e no APWeb.[4]
A orde foi recoñecida polo APG III (2009),[1] e o Linear APG III (2009)[2] asignoulle os números de familia 62 a 75. A orde xa fora recoñecida polo APG II (2003).[8]
A orde, como actualmente está circunscrita, consiste en 14 familias e aproximadamente 26 800 especies. Os límites dalgunhas familias aínda son pouco claros, e como se describiu na sección de filoxenia, algúns botánicos propuxeron que se combinen moitas das familias que aquí se recoñecen. Por exemplo, o APG III e o APWeb tratan a Alliaceae, Amaryllidaceae e Agapanthaceae, que xuntos forman un clado, como unha única familia: Amaryllidaceae sensu lato. Tamén combinan Xanthorrhoeaceae, Asphodelaceae e Hemerocallidaceae nun Xanthorrhoeaceae sensu lato, e Agavaceae, Asparagoideae, Hyacinthaceae, Ruscaceae, Themidaceae e unhas poucas familias pequenas combínanse na familia resultante chamada Asparagaceae sensu lato. Esta decisión é taxonomicamente arbitraria, pero deberían considerarse factores como a facilidade do recoñecemento e o nivel de sostén da monofilia do grupo.
As familias ou subfamilias máis representadas son Orchidaceae, Hypoxidaceae, Iridaceae, Amaryllidoideae, Allioideae, Scilloideae, Lomandroideae, Agavoideae, Asparagoideae, Convallariaceae, Asphodeloideae, e Hemerocallidoideae.
A lista de familias segundo APG III (2009),[1] xunto co número de familia segundo o Linear APG III (2009),[2] e algunhas das familias tradicionalmente recoñecidas e hoxe abandonadas, co seu eventual destino, están na lista que segue:
Sinónimos (APWeb):[4] Asparagineae J. Presl, Asphodelineae Thorne & Reveal, Hyacinthineae Link, Iridineae Engler - Agavales Hutchinson, Alliales Traub, Amaryllidales Bromhead, Asphodelales Doweld, Asteliales Dumortier, Hypoxidales Reveal & Doweld, Iridales Rafinesque, Ixiales Lindley, Narcissales Dumortier, Orchidales Rafinesque, Tecophilaeales Reveal, Xanthorrhoeales Reveal & Doweld - Iridanae Doweld, Orchidanae Doweld - Orchididae Heintze - Crinopsida Horaninov, Orchidopsida Bartling
No Angiosperm Phylogeny Website (xullo de 2008)[4] as familias son:
O sistema APG II de 2003,[8] dispón a orde Asparagales no clado das Monocotiledóneas e utiliza a seguinte circunscrición:
Nota: "+..." = Familia que opcionalmente pode segregarse da familia precedente. Deste modo, dentro deste sistema un taxónomo pode, por exemplo, situar ao xénero Hemerocallis na familia Hemerocallidaceae ou ben na familia Xanthorrhoeaceae sensu lato.
Hesperocallidaceae co seu único xénero Hesperocallis foi situado en Agavaceae polas últimas análises moleculares de ADN, por iso foi incluída en Agavaceae no APWeb.[4]
O sistema APG de 1998[7] tamén dispuxo a orde Asparagales dentro do clado das monocotiledóneas, pero utilizou unha circunscrición diferente das familias.
No volume 1998 (a,[14] b[72]) do Sistema Kubitzki utilizouse a seguinte circunscrición de familias dentro de Asparagales:
No Sistema Dahlgren (1985)[6] a orde Asparagales foi disposta na Superorde Lilianae dentro da Subclase Liliidae (= Monocotiledóneas) da Clase Magnoliopsida (= Angiospermas), e utilizou a siguiente circunscrición:
O sistema de Cronquist (1981)[55] non recoñece a orde Asparagales e dispón a moitos dos seus taxons dentro da orde Liliales (na subclase Liliidae, dentro da clase Liliopsida). Algúns xéneros estaban incluso displostos dentro da familia Liliaceae.
O Sistema Wettstein, que data de 1935, tampouco recoñecía a orde Asparagales e dispoñía a moitas das súas especies dentro da orde Liliflorales na clase ds monocotiledóneas.
O grupo troncal Asparagales data de hai uns 122 millóns de anos, o grupo coroa Asparagales de hai uns 119 millóns de anos (Janssen e Bremer 2004),[46] pero Wikström et al. (2001)[73] suxeriron idades de 107 a 98 e de 101 a 94 millóns de anos, respectivamente. Hai que ter en conta que no último estudo utilizouse unha topoloxía de Asparagales que difire substancialmente da aceptada aquí (especialmente preto da base da árbore).
A diverxencia do clado que contén todas as familias agás Hypoxidaceae sensu lato, Boryaceae e Orchidaceae, datouse en hai uns 84 millóns de anos (Eguiarte 1995).[74]
O "Núcleo dos Asparagales" sepárase de Xanthorrhoeaceae sensu lato hai uns 93 millóns de anos, a diverxencia dentro do clado comeza hai uns 91 millóns de anos (Janssen e Bremer 2004),[46] en cambio en Wikström et al. (2001)[73] estímanse datas de 61 a 54 e de 58 a 51 millóns de anos, respectivamente.
O grupo troncal Alliaceae sensu lato está datado nuns 91 millóns de anos ata o presente, a diverxencia dentro do grupo coroa Alliaceae sensu lato empeza hai uns 87 millóns de anos (Janssen e Bremer 2004).[46] Nótese que os fungos que infectan a Allium e outras aliáceas son bastante diferentes dos que se encontran en Amaryllidaceae (ver por exemplo Savile 1962).[75]
O grupo troncal Asparagaceae sensu lato está datado nuns 91 millóns de anos, a diverxencia dentro do grupo coroa Asparagaceae sensu lato empeza hai uns 89 millóns de anos (Janssen e Bremer 2004).[46] Eguiarte (1995),[74] porén, suxire que Agavaceae-Nolinaceae diverxiron hai só uns 47 millóns de anos.
Disposición das follas basais (Agapanthus).
Disposición das follas na extremidade do tronco en Yucca.
Hábito de crecemento de Rhodohypoxis.
Flor de Cattleya (Orchidaceae).
Flor de Hymenocallis (Amaryllidaceae).
Flores de Convallaria.
|data-acceso= require |url= (Axuda) |data-acceso= require |url= (Axuda) |data-acceso= require |url= (Axuda) |data-acceso= require |url= (Axuda) |data-acceso= require |url= (Axuda) |data-acceso= require |url= (Axuda) |data-acceso= require |url= (Axuda) |data-acceso= require |url= (Axuda) |data-acceso= require |url= (Axuda) |data-acceso= require |url= (Axuda) |data-acceso= require |url= (Axuda) (en chinés) |data-acceso= require |url= (Axuda) |data-acceso= require |url= (Axuda) |data-acceso= require |url= (Axuda) |data-acceso= require |url= (Axuda) |data-acceso= require |url= (Axuda) |data-acceso= require |url= (Axuda) |data-acceso= require |url= (Axuda) |data-acceso= require |url= (Axuda) |data-acceso= require |url= (Axuda) |data-acceso= require |url= (Axuda) |data-acceso= require |url= (Axuda) |data-acceso= require |url= (Axuda) |data-acceso= require |url= (Axuda) |data-acceso= require |url= (Axuda) |data-acceso= require |url= (Axuda) |data-acceso= require |url= (Axuda) |data-acceso= require |url= (Axuda) |data-acceso= require |url= (Axuda) |data-acceso= require |url= (Axuda) |data-acceso= require |url= (Axuda) Asparagales é unha orde de plantas anxiospermas utilizada en sistemas de clasificación modernos como o sistema de clasificación APG III do ano 2009 e o APWeb, que comprende polo menos a familia Asparagaceae. Asparagales é unha orden que se recoñeceu moi recentemente nos sistemas de clasificación e moitas das súas familias pertencían á antiga orde Liliales. Liliales fora unha orde moi ampla na cal se situaban todas as monocotiledóneas de tépalos vistosos e sen amidón no endosperma, que fora difícil de dividir en familias porque os seus caracteres morfolóxicos non aparecían en padróns que delimitasen claramente grupos. Co tempo empezouse a recoñecer a orde Asparagales, especialmente cando as análises moleculares de ADN indicaron que Liliales debía ser dividida polo menos en Liliales, Asparagales e Dioscoreales. Asparagales é unha orde bastante grande, consta de 14 familias e unhas 26 000 especies. Aínda que a orde está claramente circunscrita baseándose nas análises de secuencias de ADN pero é dificultosa de definir morfoloxicamente, xa que é un dos grupos de monocotiledóneas estruturalmente máis diverso. Así, a orde inclúe desde plantas herbáceas de non máis de 15 cm ata algunhas especies de Agave que superan os 10 m de altura. Nestes sentido, a súa separación da orde Liliales é particularmente problemática.
Unha das características que definen esta orde, e posible apomorfía (ademais dos datos moleculares), é a presenza de fitomelanina, un pigmento escuro presente nas cubertas das sementes. As fitomelaninas encóntranse na maioría das familias de Asparagales, pero perdéronse aparentemente nalgúns taxons, e o exemplo máis salientable nese aspecto é Orchidaceae.
A maioría das especies de Asparagales son plantas herbáceas perennes, aínda que hai algúns exemplos de plantas gabeadoras (certas especies de Asparagus, subfamilia Asparagoideae) e leñosas (algúns Aloe, subfamilia Asphodeloideae). Case todas as especies desta orde producen unha roseta apertada de follas na base das plantas (son "plantas en roseta"), outras levan esa mesma disposición de follas no extremo dun tronco (Yucca), e nuns poucos casos as follas aparecen ao longo do talo. As flores frecuentemente están no extremo do talo.
Dentro das monocotiledóneas e desde o punto de vista económico, a orde Asparagales é a segunda en importancia despois da orde Poales (a cal comprende os pastos e os cereais). A orde contén moitas especies de importancia hortícola (cebola, allo, espárrago, porro), en floricultura (Freesia, gladíolo, iris, moitas orquídeas), xardinería (Hemerocallis, Convallaria, Agapanthus) e especias (vainilla).
Desde o punto de vista da súa taxonomía, Asparagales é unha orde suxerida por Huber (1977), logo adoptado por Dahlgren et al. (1985), e reafianzada en 1998 polo Grupo de Filoxenia das Anxiospermas (en inglés orixinal, "Angiosperm Phylogeny Group", siglas APG), retomado no 2003 polo APG II, e en 2009 polo APG III), e tamén polo APWeb (2001 ata agora). Porén, as delimitacións das familias de Asparagales sufriron unha serie de mudanzas en anos recentes, e aínda se necesitan máis estudos para que se estabilice a súa circunscrición e as súas relacións.
Asparagales, biljni red iz razreda Liliopsida[1] kojemu (po jednom sistemu) pripada preko 36 600 vrsta u 14 niže navedenih porodica[2].
Asparagales adalah salah satu bangsa tumbuhan berbunga yang termasuk dalam klad Monokotil menurut Sistem klasifikasi APG II).
Dalam Sistem APG II (2003), bangsa ini mencakup banyak suku, yaitu
Catatan: "+ ..." = suku yang boleh dipisahkan dari induknya, bila diperlukan.
Ke dalam bangsa ini tercakup beberapa suku penting, seperti suku anggrek-anggrekan (Orchidaceae) dan suku bawang-bawangan (Alliaceae).
Sistem Cronquist tidak mengakui keberadaan bangsa ini.
Asparagales adalah salah satu bangsa tumbuhan berbunga yang termasuk dalam klad Monokotil menurut Sistem klasifikasi APG II).
Laukabálkur (fræðiheiti: Asparagales) er ættbálkur einkímblöðunga. Einkennisætt ættbálksins er sperglaætt (Asparagaceae) en hverjar aðrar ættir hafa verið settar í þennan ættbálk hefur verið mjög mismunandi í gegnum tíðina.
Þar sem "+ ..." merkir mögulega sérstaka ætt sem þá er klofin úr fyrrnefndri ætt.
Laukabálkur (fræðiheiti: Asparagales) er ættbálkur einkímblöðunga. Einkennisætt ættbálksins er sperglaætt (Asparagaceae) en hverjar aðrar ættir hafa verið settar í þennan ættbálk hefur verið mjög mismunandi í gegnum tíðina.
Asparagales Link è un ordine di piante monocotiledoni introdotto nel 1998 dal nuovo sistema di classificazione APG, creato dall'Angiosperm Phylogeny Group. Nel vecchio sistema di classificazione, le famiglie ora incluse nelle Asparagales erano incluse nell'ordine delle Liliales, e alcuni di questi generi erano inclusi nella famiglia delle Liliaceae, mentre le Orchidaceae erano inserite in un ordine autonomo, quello delle Orchidales. L'ordine prende il nome dal genere Asparagus.
Secondo la classificazione APG IV l'ordine Asparagales comprende le seguenti famiglie:[1]
Asparagales Link è un ordine di piante monocotiledoni introdotto nel 1998 dal nuovo sistema di classificazione APG, creato dall'Angiosperm Phylogeny Group. Nel vecchio sistema di classificazione, le famiglie ora incluse nelle Asparagales erano incluse nell'ordine delle Liliales, e alcuni di questi generi erano inclusi nella famiglia delle Liliaceae, mentre le Orchidaceae erano inserite in un ordine autonomo, quello delle Orchidales. L'ordine prende il nome dal genere Asparagus.
Asparagales sunt ordo plantarum florentium. Ordo familiam semper comprehendit Asparagacearum, sed alia classificationis systemata alias familias diverse amplectuntur. Hic plantarum grex se evoluisse inter Aevum Cretaceum habetur.
Systema APG II anni 2003 hunc ordinem in clado monocotyledonum ponit, et sequentem circumscriptionem adhibet:
Nota: "+ ..." = segregata familia opinabilis, quae potest removeri a priore familia.
Systema APG anni 1998 ordinem etiam in clado monocotyledonum posuit, sed hanc circumscriptionem adhibuit:
Volumen anni 1998 in serie Kubitzkiana librorum de plantis vascularibus hanc circumscriptionem adhibuit:
Systema Dahlgrenianum ordinem in superordine Lilianarum in subclassi Liliidarum (= monocotyledonum) classis Magnoliopsidorum (= angiospermarum) et hanc circumscriptionem adhibuit:
Asparagales sunt ordo plantarum florentium. Ordo familiam semper comprehendit Asparagacearum, sed alia classificationis systemata alias familias diverse amplectuntur. Hic plantarum grex se evoluisse inter Aevum Cretaceum habetur.
Smidriečiai (lot. Asparagales, vok. Orchideen) – magnolijūnų (Manoliophyta) skyriaus lelijainių (Liliopsida) klasės augalų eilė.
Smidriečiai (lot. Asparagales, vok. Orchideen) – magnolijūnų (Manoliophyta) skyriaus lelijainių (Liliopsida) klasės augalų eilė.
Asparagales (Nederlands: aspergeachtigen) is de botanische naam van een orde van eenzaadlobbige planten. Een orde onder deze naam werd niet erkend in het Cronquist systeem (1981), maar wel in het APG-systeem (1998), het APG II-systeem (2003) en het APG III-systeem (2009). Samen met de ordes Liliales en Poales vormen ze in de Europese flora de belangrijkste ordes van eenzaadlobbigen. Er zijn geen eenvoudig herkenbare eigenschappen. Bij de Asparagales komen parasiterende planten voor, bij de Liliales niet. De Nederlandse naam voor deze orde is Aspergeorde.
De indeling volgens het APG III-systeem (2009) is als volgt:
Hierbij zijn de families tussen "[+ ... ]" optioneel: deze families kunnen al dan niet afgesplitst worden van de voorgaande familie. Dit betekent dat voor de families Alliaceae, Asparagaceae en Xanthorrhoeaceae ieder twee verschillende omschrijvingen toegestaan zijn.
Dit is een wijziging ten opzichte van het APG-systeem (1998), waar de samenstelling de volgende was:
Asparagales er en orden av einfrøblada plantar. Dei fleste artane i ordenen høyrer til i orkidéfamilien, som kanskje er den største familien av blomsterplantar, men nokre av dei andre familiane er også artsrike. Asparagales omfattar omtrent halvparten av alle einfrøblada planter og 10–15 % av alle blomsterplantar. Ordenen omfattar fleire kulturelt og økonomisk viktige artar, som smaksgjevarane lauk, kvitlauk, purre, asparges og vanilje, og prydblomar som Freesia, Gladiolus, Iris, orkidear, dagliljer, liljekonvall og påskeliljefamilien.
I eldre systematikk, slik som Cronquist-systemet, blei desse plantene rekna til Liliales, og nokre var med i den store liljefamilien (Liliaceae). Molekylærgenetikk viser at Asparagales er ei naturleg gruppe, men gruppa er vanskeleg å definera ut frå morfologi. Dei fleste artane i ordenen er fleirårige urteplantar (staudar), men nokre er klatreplantar og andre veks meir som tre. Mange plantar har også organ til å lagar næring i, som lauk, jordstengel eller rotknoll. Blada dannar som regel ein bladrosett, gjerne heilt nede ved bakken eller heilt i toppen, men ein del har også blad langs stengelen. Blomane er «liljeaktige» med seks blomeblad og opptil seks pollenberarar.
Plantane i ordenen skilde seg truleg ut frå andre einfrøbladingar for rundt 120–130 millionar å sidan, tidleg i krittida. Problem med å klassifisera familiane det er snakk om gjer at slike estimat er usikre.
Det har vore mange oppfatningar om systematikken i gruppa. Denne artikkelen følgjer APG IV-systemet med 14 familiar. Aloefamilien (Asphodelaceae), laukfamilien (Amaryllidaceae) og aspargesfamilien (Asparagaceae) er utvida ved at fleire tidlegare familiar no har status som underfamiliar. Orkidéfamilien (Orchidaceae) er systergruppe til resten av Asparagales.
Kladogrammet under viser slektskapa ein meiner finst innanfor Asparagales. Norske namn følgjer Artsdatabanken.
Asparagalesorkidéfamilien (Orchidaceae)
sverdliljefamilien (Iridaceae)
Hemerocallidoideae (= Hemerocallidaceae, «dagliljefamilien»)
Xanthorrhoeoideae (= Xanthorrhoeaceae s.s.)
Asphodeloideae (= Asphodelaceae,
«aloefamilien» i snever tyding)
Agapanthoideae (= Agapanthaceae)
Allioideae (= Alliaceae s.s.,
«laukfamilien» i snever tyding)
Amaryllidoideae (= Amaryllidaceae s.s.,
«narsissfamilien»)
Aphyllanthoideae (= Aphyllanthaceae)
Brodiaeoideae (= Themidaceae)
Scilloideae (= Hyacinthaceae,
«hyasintfamilien»)
Agavoideae (= Agavaceae, «agavefamilien»)
Lomandroideae (= Laxmanniaceae)
Asparagoideae (= Asparagaceae s.s.,
«aspargesfamilien» i snever tyding)
Nolinoideae (= Ruscaceae)
Asparagales er en orden av einfrøblada plantar. Dei fleste artane i ordenen høyrer til i orkidéfamilien, som kanskje er den største familien av blomsterplantar, men nokre av dei andre familiane er også artsrike. Asparagales omfattar omtrent halvparten av alle einfrøblada planter og 10–15 % av alle blomsterplantar. Ordenen omfattar fleire kulturelt og økonomisk viktige artar, som smaksgjevarane lauk, kvitlauk, purre, asparges og vanilje, og prydblomar som Freesia, Gladiolus, Iris, orkidear, dagliljer, liljekonvall og påskeliljefamilien.
I eldre systematikk, slik som Cronquist-systemet, blei desse plantene rekna til Liliales, og nokre var med i den store liljefamilien (Liliaceae). Molekylærgenetikk viser at Asparagales er ei naturleg gruppe, men gruppa er vanskeleg å definera ut frå morfologi. Dei fleste artane i ordenen er fleirårige urteplantar (staudar), men nokre er klatreplantar og andre veks meir som tre. Mange plantar har også organ til å lagar næring i, som lauk, jordstengel eller rotknoll. Blada dannar som regel ein bladrosett, gjerne heilt nede ved bakken eller heilt i toppen, men ein del har også blad langs stengelen. Blomane er «liljeaktige» med seks blomeblad og opptil seks pollenberarar.
Plantane i ordenen skilde seg truleg ut frå andre einfrøbladingar for rundt 120–130 millionar å sidan, tidleg i krittida. Problem med å klassifisera familiane det er snakk om gjer at slike estimat er usikre.
Det har vore mange oppfatningar om systematikken i gruppa. Denne artikkelen følgjer APG IV-systemet med 14 familiar. Aloefamilien (Asphodelaceae), laukfamilien (Amaryllidaceae) og aspargesfamilien (Asparagaceae) er utvida ved at fleire tidlegare familiar no har status som underfamiliar. Orkidéfamilien (Orchidaceae) er systergruppe til resten av Asparagales.
Asparagales er en orden av enfrøbladete planter. De fleste artene inngår i orkidéfamilien, som kanskje er den største familien av blomsterplanter, men noen av de andre familiene er også artsrike. Asparagales omfatter omtrent halvparten av alle enfrøbladete planter og 10–15 % av alle blomsterplanter.
I eldre systematikk, for eksempel Cronquist-systemet, ble disse plantene regnet til Liliales, og noen tilhørte den store liljefamilien (Liliaceae). Molekylærgenetikk viser at Asparagales er en naturlig gruppe, men gruppen er vanskelig å definere utfra morfologi.
Det har vært mange oppfatninger om systematikken innenfor gruppen. Denne artikkelen følger APG IV-systemet med 14 familier. Aloefamilien (Asphodelaceae), løkfamilien (Amaryllidaceae) og aspargesfamilien (Asparagaceae) er utvidet ved at flere tidligere familier nå har status som underfamilier. Orkidéfamilien (Orchidaceae) er søstergruppen til resten av Asparagales.
Følgende kladogram viser antatt slektskap innenfor Asparagales. Norske navn følger Artsdatabanken.
Asparagalesorkidéfamilien (Orchidaceae)
sverdliljefamilien (Iridaceae)
Hemerocallidoideae (= Hemerocallidaceae, «dagliljefamilien»)
Xanthorrhoeoideae (= Xanthorrhoeaceae s.s.)
Asphodeloideae (= Asphodelaceae,
«aloefamilien» i snever betydning)
Agapanthoideae (= Agapanthaceae)
Allioideae (= Alliaceae s.s.,
«løkfamilien» i snever betydning)
Amaryllidoideae (= Amaryllidaceae s.s.,
«narsissfamilien»)
Aphyllanthoideae (= Aphyllanthaceae)
Brodiaeoideae (= Themidaceae)
Scilloideae (= Hyacinthaceae,
«hyasintfamilien»)
Agavoideae (= Agavaceae, «agavefamilien»)
Lomandroideae (= Laxmanniaceae)
Asparagoideae (= Asparagaceae s.s.,
«aspargesfamilien» i snever betydning)
Nolinoideae (= Ruscaceae)
Asparagales er en orden av enfrøbladete planter. De fleste artene inngår i orkidéfamilien, som kanskje er den største familien av blomsterplanter, men noen av de andre familiene er også artsrike. Asparagales omfatter omtrent halvparten av alle enfrøbladete planter og 10–15 % av alle blomsterplanter.
I eldre systematikk, for eksempel Cronquist-systemet, ble disse plantene regnet til Liliales, og noen tilhørte den store liljefamilien (Liliaceae). Molekylærgenetikk viser at Asparagales er en naturlig gruppe, men gruppen er vanskelig å definere utfra morfologi.
Det har vært mange oppfatninger om systematikken innenfor gruppen. Denne artikkelen følger APG IV-systemet med 14 familier. Aloefamilien (Asphodelaceae), løkfamilien (Amaryllidaceae) og aspargesfamilien (Asparagaceae) er utvidet ved at flere tidligere familier nå har status som underfamilier. Orkidéfamilien (Orchidaceae) er søstergruppen til resten av Asparagales.
Szparagowce (Asparagales Bromhead) – rząd roślin należących do klasy jednoliściennych. Wyróżnia się w jego obrębie 14 rodzin z ponad 1,1 tysiącem rodzajów i ponad 36,2 tysiące gatunków, z czego 26 tysięcy należy do rodziny storczykowatych[1]. Należące tu rośliny to często geofity z owocami typu torebka i jagoda[2]. Szparagowce spotykane są na całym świecie, ale do obszarów o ich szczególnym zróżnicowaniu należy południowa Afryka (choć akurat storczykowatych tu jest niewiele)[1].
W dawniejszych systemach (np. Cronquista z 1988 r.) rząd ten nie pojawia się wcale, a przedstawiciele tej linii rozwojowej zaliczani byli zwykle do parafiletycznego w ówczesnym ujęciu rzędu liliowców (Liliales). Późniejsze badania biochemiczne w zakresie serologicznych reakcji białek nasion i cytologiczne (obecność komórek śluzowych i rafidów) pozwoliły na wyodrębnienie tej grupy roślin w randze rzędu (system Reveala (1999) i Takhtajana (1997)). W końcu badania molekularne i prace taksonomów z Angiosperm Phylogeny Group spowodowały wydzielenie w systemie APG II (2003) dużego kladu oznaczonego jako rząd Asparagales. Obejmuje on w tym ujęciu grupę roślin, w obrębie której najstarszą linią rozwojową są storczykowate (Orchidaceae). W systemie APG III (2009) podział rzędu został uproszczony, poprzez scalenie szeregu rodzin w taksony o szerszym ujęciu[3]. Rodzina żółtakowate (Xanthorrhoeaceae) została powiększona o wcześniej wyodrębniane w randze rodzin złotogłowowe (Asphodeloideae) i liliowcowe (Hemerocallidoideae). Rodzina amarylkowate (Amaryllidaceae) została powiększona o agapantowe (Agapanthoideae) i czosnkowe (Allioideae). W końcu szparagowate (Asparagaceae) objęły w sumie 7 taksonów wcześniej wyodrębnianych jako osobne rodziny[4]. W systemie APG IV z 2016 wprowadzono tylko korektę w nomenklaturze zmieniając nazwę rodziny żółtakowate Xanthorrhoeaceae na złotogłowowate Asphodelaceae[5].
tatarakowce Acorales
żabieńcowce Alismatales
pochrzynowce Dioscoreales
pandanowce Pandanales
liliowce Liliales
szparagowce Asparagales
arekowce Arecales
wiechlinowce Poales
komelinowce Commelinales
imbirowce Zingiberales
storczykowate Orchidaceae
asteliowate Asteliaceae
przyklękowate Hypoxidaceae
kosaćcowate Iridaceae
złotogłowowate Asphodelaceae
amarylkowate Amaryllidaceae
szparagowate Asparagaceae
W systemie Reveala (2007) rząd szparagowców nie jest wyróżniany. Klad, którego na kladogramie według APW bazową linią jest rodzina Ixioliriaceae, określony jest przez Reveala jako rząd Iridales Raf. Podział tego kladu na rodziny też różni się nieco od ujęcia prezentowanego na APW (wydzielonych jest kilka drobnych rodzin, łączonych przez APW). Starsze linie rozwojowe kladu szparagowców według APW zaliczone zostały przez Reveala do rzędu storyczykowców Orchidales Raf.[6].
Gromada okrytonasienne (Magnoliophyta Cronquist), podgromada Magnoliophytina Frohne & U. Jensen ex Reveal, klasa jednoliścienne (Liliopsida Brongn.), podklasa liliowe (Liliidae J.H. Schaffn.), nadrząd Lilianae Takht., rząd szparagowce Asparagales Bromhead[7].
Szparagowce (Asparagales Bromhead) – rząd roślin należących do klasy jednoliściennych. Wyróżnia się w jego obrębie 14 rodzin z ponad 1,1 tysiącem rodzajów i ponad 36,2 tysiące gatunków, z czego 26 tysięcy należy do rodziny storczykowatych. Należące tu rośliny to często geofity z owocami typu torebka i jagoda. Szparagowce spotykane są na całym świecie, ale do obszarów o ich szczególnym zróżnicowaniu należy południowa Afryka (choć akurat storczykowatych tu jest niewiele).
Aspargales é uma ordem de plantas com flor que na circunscrição taxonómica que lhe é dada pelo sistema de classificação APG IV (2016) é a maior das ordens de monocotiledóneas, agrupando 14 famílias, com 1122 géneros e cerca de 36 000 espécies, entre as quais algumas com elevado interesse económico, como os espargos, as cebolas, o sisal, a baunilha e múltiplas plantas ornamentais.[4] Apesar de claramente circunscrita com base na sua filogenia molecular, a ordem é difícil de definir morfologicamente, pois os seus membros são estruturalmente muito diversos, indo dos pequenos caméfitos às lianas e a alguns arbustos e pequenas árvores, embora contenha maioritariamente espécies herbáceas perenes com elevada proporção de geófitos.
A ordem Asparagales, cujo nome deriva do táxon tipo deste agrupamento taxonómico, a família Asparagaceae, agrupa as monocotiledóneas lilioides conhecidas por «liliáceas asparagoides» nos modernos sistemas de classificação, tais como o Angiosperm Phylogeny Group (APG) e o Angiosperm Phylogeny Web.
Esta ordem apenas recentemente foi reconhecida nos sistemas de classificação, proposta pela primeira vez pelo botânico alemão Herbert Huber em 1977 e posteriormente adoptado no sistema de Dahlgren de 1985 e, em seguida, no sistema de classificação APG I de 1998 (e mantida nas subsequente versões II, II e IV, de 2003, 2009 e 2016, respectivamente). Antes disso, muitas das famílias que agora a integram estavam atribuídas à antiga ordem Liliales, uma ordem muito grande, que continha contendo quase todas as monocotiledóneas com tépalas coloridas e sem amido no endosperma das suas sementes. Contudo, a análise de sequências de DNA indicou que muitos dos táxons previamente incluídos na ordem Liliales deveriam na verdade ser redistribuídos em três ordens, Liliales, Asparagales e Dioscoreales.
A circunscrição taxonómica da ordem Asparagales, e das suas famílias constituintes, sofreu uma série de mudanças nos últimos anos e resultados de futuros estudos podem levar a mais mudanças, mas também, em última instância, a uma maior estabilidade. Na circunscrição adoptada pelo sistema de classificação APG IV, de 2016, a ordem Asparagales é a maior ordem de monocotiledóneas, com 14 famílias, 1122 géneros e cerca de 36 000 espécies.
A ordem está claramente circunscrita com base na filogenética molecular, mas é difícil de definir morfologicamente, uma vez que os seus membros são estruturalmente muito diversos. A maioria das espécies de Asparagales são herbáceas perenes, embora ocorram trepadeiras (ou lianas) e alguns sejam semelhantes a árvores. A ordem contém muitos geófitos (com bolbos, cormos e vários tipos de tubérculos). De acordo com a sequência telomérica, pelo menos dois pontos de mudança evolutivos aconteceram dentro da ordem. A sequência basal é formada por TTTAGGG, como na maioria das plantas superiores, mas o motivo basal foi alterado para TTAGGG, semelhante aos vertebrados e, finalmente, o motivo mais divergente CTCGGTTATGGG aparece em Allium. Uma das características definidoras (sinapomorfias) da ordem é a presença de fitomelanina, um pigmento preto presente no tegumento das sementes, criando uma crosta escura. A fitomelanina é encontrada na maioria das famílias de Asparagales (embora não em Orchidaceae, família considerada o grupo-irmão do resto da ordem).
As folhas de quase todas as espécies formam uma roseta apertada, seja na base da planta seja no final da haste da planta, mas ocasionalmente ao longo do caule. As flores não são particularmente distintas, sendo do tipo lírio, com seis tépalas e até seis estames.
Acredita-se que a ordem tenha primeiro divergido das outras monocotiledóneas com as quais é aparentada há cerca de 120-130 milhões de anos atrás (no início do Cretáceo), embora dada a dificuldade em classificar as famílias envolvidas, as estimativas sejam provavelmente incertas.
Do ponto de vista económico, a ordem Asparagales é a segunda em importância dentro das monocotiledóneas, depois da ordem Poales (que inclui gramíneas e cereais). As espécies são usadas como alimentos e condimentos (por exemplo, cebola, alho, alho-porro, aspargos, baunilha, açafrão), em aplicações em medicamentos ou cosméticos (Aloe), como flores de corte (por exemplo, frésia, gladíolo, íris, orquídeas), e como ornamentais em jardins e paisagismo (por exemplo, lírios, lírio-do-vale e Agapanthus).
Embora a maioria das espécies incluídas na ordem sejam pequenas herbáceas, algumas com menos de 15 cm de altura, há algumas trepadeiras (entre as quais algumas espécies de Asparagus), bem como vários géneros arborescentes (como Agave, Cordyline, Yucca, Dracaena e Aloe), que podem exceder os 10 m de altura. Os géneros suculentos ocorrem em várias famílias (como o género Aloe).
Embora em alguns casos, as folhas sejam produzidas ao longo do caule, quase todas as espécies apresentam uma filotaxia característica, com um padrão de distribuição das folhas em aglomerado compacto que dá origem à formação de uma roseta) localizada na base da planta ou no extremo dos caules, como ocorrem com o género Yucca. As flores, frequentemente agrupadas no final do caule principal da planta, não são em geral particularmente distintas, sendo predominantemente do 'tipo lírio', com seis tépalas, livres ou fundidas a partir da base, e até seis estames.
As Asparagales são geralmente diferenciados das Liliales pela falta de marcações nas tépalas, a presença de nectários septais nos ovários, em vez de na base das tépalas ou filamentos dos estames, e a presença de crescimento secundário. Geralmente são geófitos, mas com folhas lineares e falta de venação reticular fina.
As sementes apresentam caracteristicamente a epiderme externa obliterada (na maioria das espécies com frutos carnosos) ou, se presentes, têm uma camada de fitomelanina de coloração preto carbonáceo nas espécies com frutos secos. A parte interna do tegumento é geralmente colapsada, ao contrário dos Liliales cujas sementes têm epiderme externa bem desenvolvida, carecem de fitomelanina e geralmente apresentam uma camada interna celular.
As ordens que foram separadas da antiga circunscrição da ordem Liliales são difíceis de caracterizar, já que nenhuma característica morfológica única parece ser diagnóstico seguro da ordem Asparagales:
Como circunscrito pelo Angiosperm Phylogeny Group no sistema de classificação APG IV, as Asparagales são a mais numerosa e diversa das ordens em que estão subdivididas as monocotiledóneas («monocots», Monocotyledoneae ou Liliopsida), com 14 famílias, 1122 géneros e entre 25 000 a 42 000 espécies, o que representa cerca de 50% de todas as monocots e 10–15% da totalidade das plantas com flor (Angiospermae).[9][10]
A atribuição de autoridade botânica para o nome Asparagales pertence a Johann Heinrich Friedrich Link (1767-1851) que cunhou a palavra «Asparaginae» em 1829 para um táxon de ordem superior que incluía o género Asparagus,[11] embora Michel Adanson e Antoine Laurent de Jussieu também o tivessem feito antes. As circunscrições anteriores de Asparagales atribuíram o nome a Edward Bromhead (1838), que foi o primeiro a usar o termo «Asparagales».[3]
O género tipo, Asparagus, a partir do qual o nome da ordem é derivado, foi descrito por Carl Linnaeus em 1753, com 10 espécies.[12] No seu sistema de classificação, Lineu colocou o género Asparagus entre as Hexandria Monogynia (seis estames, um carpelo) da classificação sexual que constitui a estrutura da sua obra Species Plantarum.[13]
A maioria dos taxa que agora são considerados como parte das Asparagales foi historicamente colocada dentro de uma família Liliaceae muito grande e diversa, com uma circunscrição bem mais alargada que a presentemente considerada como filogeneticamente correcta. A família Liliaceae foi descrita pela primeira vez por Michel Adanson em 1763,[14] que no seu esquema taxonómico criou oito secções, incluindo a secção «Asparagi» with Asparagus e três outros géneros.[15]
O sistema de organização dos géneros em famílias é geralmente creditado a Antoine Laurent de Jussieu, que descreveu formalmente os Liliaceae e a família-tipo de Asparagales, as Asparagaceae, como «Lilia» e «Asparagi», respectivamente, em 1789.[16] Jussieu estabeleceu o sistema hierárquico que está na base da moderna taxonomia (filogenia), colocando o género Asparagus e géneros próximo no interior da divisão das Monocotyledon, uma classe (III) das Stamina Perigynia,[17] e na 'ordem' (ordo) Asparagi, dividida em três subfamílias.[18] O uso do termo Ordo (ordem) naquela época estava mais perto do que agora se entende por família, ao invés do actual conceito de ordem.[19][20]
Ao criar o seu sistema de classificação, Jussieu usou uma forma modificada da «classificação sexual de Linnaeus», que, em vez de se basear exclusivamente no número de estames e carpelos, usava também as posições relativas destes órgãos na estrutura floral. Enquanto o agrupamento taxonómico Stamina Perigynia de Jussieu também incluía um conjunto de 'ordens' que depois dariam eventualmente origem às famílias que presentemente integram as Asparagales, tais como as «Asphodeli» (Asphodelaceae), «Narcissi» (Amaryllidaceae) e «Irides» (Iridaceae), estando as restantes presentemente alocadas a outras ordens. As «Asparagi» de Jussieu passaram pouco depois a ser referidas como as Asparagacées na literatura em língua francesa (em latim: Asparagaceae).[21] Entretanto, as 'Narcissi' passaram a ser designadas por 'Amaryllidées' (Amaryllideae) em 1805, por Jean Henri Jaume Saint-Hilaire, usando Amaryllis como a espécie tipo em vez de Narcissus, e em consequência passou a ser-lhe atribuída a autoridade taxonómica para a família Amaryllidaceae.[22] Em 1810 Robert Brown propôs que um subgrupo de Liliaceae fosse distinguido com base na posição do ovário, designando-o por Amaryllideae,[23] e em 1813 de Candolle descreveu as «Liliacées Juss.» e «Amaryllidées Brown» como duas famílias autónomas e bem distintas.[24]
A literatura sobre a organização de géneros em famílias e a sua organização nos níveis taxonómicos superiores tornou-se disponível na língua inglesa com a publicação da obra de Samuel Frederick Gray intitulada A natural arrangement of British plants (1821).[25] Gray usou uma combinação da «classificação sexual» de Linnaeus com a «classificação natural» de Jussieu para agrupar várias famílias que tinham em comum seis estames iguais, um único estilo e um perianto que era simples e petalóide, mas não usava nomes formais para essas classificações de nível mais elevado. Dentro do grupo, separou as famílias pelas características dos seus frutos e sementes e tratou grupos de géneros com essas características em comum como famílias separadas, como Amaryllideae, Liliaceae, Asphodeleae e Asparageae.[26]
A circunscrição de Asparagales tem sido uma fonte de dificuldade para muitos botânicos desde a época de John Lindley (1846), o outro taxonomista britânico importante do início do século XIX. No seu sistema de classificação, o sistema de Lindley, publicado na sua obra intitulada An Introduction to the Natural System of Botany (1830),[27] seguiu parcialmente o sistema de Jussieu ao descrever uma subclasse que designou por «Endogenae», ou «Plantas Monocotiledóneas» (preservando a designação Endogenæ phanerogamæ criada por de Candolle),[28] dividida em duas tribos, Petaloidea e Glumaceae. Subdividiu a primeiro daquelas tribos, frequentemente conhecido como «monocotiledóneas petalóides», em 32 ordens, incluindo, para além das Liliaceae (definidas estritamente), a maioria das famílias consideradas como constituintes da actual ordem Asparagales como é presentemente circunscrita, incluindo as Amaryllideae.
Em 1846, na última versão do seu sistema de classificação,[29] John Lindley expandiu e refinou muito o tratamento das monocotiledóneas, introduzindo uma classificação intermediária («Alliances» ou seja, «alianças») e tribos dentro das ordens (ou seja das actuais famílias). Neste arranjo, John Lindley colocou as Liliaceae dentro das Liliales, mas considerou-as como uma família parafilética (um "grupo residual"), onde agrupou todas Liliales não incluídas nas outras ordens, manifestando a esperança que o futuro revelasse alguma característica que permitisse agrupar melhor estas espécies. Em resultado, a ordem Liliales ficou muito grande e incluía quase todas as monocotiledóneas com tépalas coloridas e sem amido no endosperma (as monocotiledóneas lilioides). Era difícil dividir as Liliales em famílias porque os caracteres morfológicos não estavam presentes em padrões que demarcassem claramente os grupos. Essa ausência de padrões morfológicos distintivos emtre grupos manteve as Liliaceae separadas das Amaryllidaceae (Narcissales). Estas Liliaceae[30] foram divididas em 11 tribos (com 133 géneros) e as Amaryllidaceae[31] em 4 tribos (com 68 géneros), embora ambos contivessem muitos géneros que eventualmente segregariam nas ordens contemporâneas uns dos outros (Liliales e Asparagales, respectivamente). As Liliaceae seriam reduzidas a um pequeno 'núcleo' representado pela tribo Tulipae, enquanto os grandes grupos, como Scilleae e Asparagae, se tornariam parte de Asparagales como parte das Amaryllidaceae ou como famílias separadas. Enquanto das Amaryllidaceae, as Agaveae seriam parte das Asparagaceae, mas as Alstroemeriae seriam depois consideradas uma família dentro das Liliales.
O número de géneros e espécies conhecidos continuou a crescer até ao surgimento do sistema de Bentham & Hooker, de 1883, publicado em latim, várias das outras famílias que constavam do sistema de John Lindley foram absorvidas pelas Liliaceae.[32] Nesta nova classificação usaram o termo 'série' para indicar a classificação supra-família, com sete séries de monocotiledóneas (incluindo as Glumaceae), mas não usaram as designações de Lindley. No entanto, colocaram os géneros liliáceos e amaralidáceos em séries separadas. As Liliaceae[33] foram colocadas na série Coronariae, enquanto as Amaryllideae[34] foram colocados na série Epigynae. As Liliaceae passaram a ser divididas em 20 tribos (incluindo Tulipeae, Scilleae e Asparageae), e as Amaryllideae por 5 tribos (incluindo Agaveae e Alstroemerieae). Um acréscimo importante ao tratamento dado às Liliaceae foi o reconhecimento de Allieae como uma tribo distinta que acabaria por chegar às Asparagales como a subfamília Allioideae das Amaryllidaceae.[35]
O aparecimento da Origem das Espécies de Charles Darwin em 1859 mudou a maneira como os taxonomistas consideravam a classificação das plantas, levantando a questão da incorporação de informação evolutiva nos esquemas de classificação. A abordagem darwiniana conduziu ao conceito de filogenia (com o surgimento de uma estrutura lógica estrutura semelhante a uma árvore) na montagem de sistemas de classificação, começando com o sistema de Eichler,[36] poroposto por August Eichler, que estabeleceu um sistema hierárquico no qual as plantas com flores (angiospermas) foram divididas em monocotiledóneas e dicotiledóneas, posteriormente subdividindo as primeiras em sete ordens distintas. Dentro das Liliiflorae havia sete famílias, incluindo Liliaceae e Amaryllidaceae. Liliaceae incluía Allium e Ornithogalum (as modernas Allioideae) e Asparagus.[37]
Alguns anos depois, Adolf Engler, com o seu Sistema de Engler, desenvolveu as ideias de Eichler num esquema muito mais elaborado que expôs numa série de trabalhos, incluindo Die Natürlichen Pflanzenfamilien (saído em 1888 em co-autoria com Karl Anton Prantl)[38] e o Syllabus der Pflanzenfamilien (1892–1924).[39] No seu tratamento de Liliiflorae, as Liliineae eram uma subordem que incluía as famílias Liliaceae e Amaryllidaceae. As Liliaceae[40] incluíam oito subfamílias e as Amaryllidaceae[41] quatro. Neste rearranjo das Liliaceae, com menos subdivisões, as Liliales centrais foram consideradas como constituindo a subfamília Lilioideae (com Tulipae e Scilleae como tribos) e as Asparagae como Asparagoideae, enquanto as Allioideae eram preservadas agrupando os géneros aliáceos. As Allieae, Agapantheae e Gilliesieae eram as três tribos incluída nesta subfamília.[42] Nas Amaryllidacea houve poucas mudanças em relação ao sistema de Bentham & Hooker.
Uma abordagem semelhante foi adoptada por Richard Wettstein,[43] ao propor um novo sistema de classificação, que ficaria conhecido por sistema de Wettstein (1901–1935), colocou muitos dos taxa das antigas Liliales numa ordem que designou por 'Liliiflorae'.[44]
A partir do sistema de Wettstein, o botânico neerlandês Johannes Paulus Lotsy (1911) propôs dividir as Liliiflorae, incluindo as Asparagaceae, em várias famílias menores, mais homogéneas, [45] como forma de criar grupos que hoje seriam designados por monofiléticos. A proposta de Johannes Lotsy foi alargada e consolidada por Herbert Huber (1969, 1977), que propôs a divisão das Liliiflorae em quatro grupos, incluindo as Liliiflorae 'asparagoides'.[46][47]
O aparecimento do sistema de Cronquist (1968–1988), amplamente utilizado, manteve uma grande ordem Liliales circunscrita de forma muito ampla.[48][49][50]
Essas várias propostas visando separar pequenos grupos de géneros em famílias mais homogéneas tiveram pouco impacte até ao aparecimento do sistema de Dahlgren, proposto em 1985 pelo botânico sueco Rolf Dahlgren, incorporando novas informações e conceitos de carácter filogenético, incluindo a procura de sinapomorfias. Dahlgren aprofundou as ideias de Huber e popularizou-as, incluindo no seu sistema uma grande desconstrução das famílias existentes em unidades menores. Com esse objectivo propôs uma nova ordem, designado-a por Asparagales. Este foi uma das cinco ordens propostas dentro da superordem Liliiflorae. Onde Cronquist vira uma família, Dahlgren viu quarenta, distribuídas em três ordens (predominantemente Liliales e Asparagales).[51][52]
Ao longo da década de 1980, no contexto de uma revisão mais geral da classificação das angiospermas, as Liliaceae foram submetidas a um escrutínio mais intenso. No final daquela década, o Royal Botanic Gardens at Kew, o British Museum of Natural History e o Edinburgh Botanical Gardens formaram uma comissão especializada para examinar a possibilidade de, pelo menos, separar as famílias para a organização dos seus herbários. Esse comitê recomendou que fossem criadas 24 novas famílias no lugar da amplíssima família Liliaceae original, em grande parte elevando as subfamílias à categoria de famílias separadas.[53]
Dada a diversidade morfológica das espécies que a integram, a ordem Asparagales, como atualmente circunscrita, só resultado do advento da filogenética foi recentemente reconhecida nos sistemas de classificação. Na década de 1990 ocorreu um progresso considerável no conhecimento da filogenia das plantas e na teoria filogenética, permitindo a construção de uma árvore filogenética abrangendo todas as plantas com flor. O estabelecimento de novos clados representativos exigiu o abandono das classificações antigas, mas ainda amplamente utilizadas, como o sistema de Cronquist e o sistema de Thorne, baseadas principalmente na morfologia, em vez de nos dados genéticos. Esta complexa discussão acerca da evolução das plantas levou a uma grande reestruturação conceptual e a novas formas de agrupar as monocotiledóneas.[54]
O sequenciamento do gene rbcL e a análise cladística das monocotiledóneas conduziu a uma profunda redefinição das Liliales em 1995,[55][56] passando de quatro ordens morfológicas sensu Rolf Dahlgren. Tal levou à circunscrição de um grande clado representativo das Liliaceae com múltiplas espécies, todas previamente incluídas em Liliales, mas incluindo também as Calochortaceae e Liliaceae sensu Minoru N. Tamura. Esta família redefinida, que passou a ser designada por «Liliales nucleares», correspondia já à circunscrição emergente do Angiosperm Phylogeny Group (1998).[57]
A partir da revisão de 2009, correspondente ao sistema APG III, o Angiosperm Phylogeny Group colocou a ordem Asparagales, como tal, no clado das monocots,[58] já que desde o surgimento do sistema de Dahlgren, em 1985, sucessivos estudos baseados principalmente na morfologia identificaram os Asparagales como um grupo distinto, embora, por razões morfológicas, também incluíssem grupos agora filogeneticamente localizados em Liliales, Pandanales e Zingiberales.[59] Investigação filogenética mais recente veio comprovar a monofilia de Asparagales, com base na morfologia, na sequência 18S do rDNA e em outras sequências de DNA,[60][61][62][63][64] embora algumas reconstruções filogenéticas baseadas em dados moleculares tenham sugerido que Asparagales podem ser parafiléticas, com as Orchidaceae separadas dos restantes agrupamentos.[65] Esses mesmos estudos também permitiram comprovar que, no conjunto das monocotiledéneas, Asparagales é o grupo irmão do clado das comelinídeas.[54]
O cladograma que se segue mostra a colocação de Asparagales no contexto das ordens de Lilianae sensu Chase & Reveal (monocotiledóneas) com base em evidências filogenéticas moleculares.[66][58][67][68] As ordens pertencentes ao grupo das monocotiledóneas lilioides estão entre colchetes, a saber Petrosaviales, Dioscoreales, Pandanales, Liliales e Asparagales.[69] Esses agrupamentos taxonómicos constituem um grupo parafilético, isto é, são um grupo com um ancestral comum que não inclui todos os descendentes directos (neste caso as commelinídeas como o grupo irmão de Asparagales). Para formar um clado, todos os grupos unidos por linhas grossas precisariam ser incluídos. Enquanto as Acorales e Alismatales são colectivamente referidas como "monocotiledóneas alismatídeas" (monocotiledóneas basais ou de ramificação inicial), os clados restantes (monocotiledóneas lilioides e commelinídeas) são em geral referidas como "monocotiledóneas nucleares".[70] A relação entre estas ordens (com excepção das duas que constituem grupos-irmão) é a presença de pectinatos, característica que marca a divergência na sucessão ao longo da linha que leva às comelinídeas.[67] Os números constantes do cladograma indicam, para cada grupo coroa (ancestral comum mais recente com cada uma das espécies do clado), os tempos estimados de divergência em milhões de anos atrás.[68] Tendo em consideração essas relações, a ordem Asparagales insere-se nas Monocots da seguinte forma:[71][72]
Lilianae sensu Chase & Reveal (monocots) 131[66]Dioscoreales 115
Pandanales 91
Liliales 121
Asparagales 120
Simplificando, o grupo taxonómico onde se inserem as Asparagales pode ser respresentado pelo seguinte cladograma:
MonocotsAsparagales
Uma árvore filogenética para os Asparagales, geralmente desagregada até ao nível de família, mas incluindo grupos que embora reduzidos à categoria de subfamília foram até recentemente amplamente tratados como famílias, leva ao seguinte cladograma:[9][1]
AsparagalesHemerocallidoideae (= Hemerocallidaceae)
Xanthorrhoeoideae (= Xanthorrhoeaceae s.s.)
Asphodeloideae (= Asphodelaceae)
Agapanthoideae (= Agapanthaceae)
Allioideae (= Alliaceae s.s.)
Amaryllidoideae (= Amaryllidaceae s.s.)
Aphyllanthoideae (= Aphyllanthaceae)
Agavoideae (= Agavaceae)
Lomandroideae (= Laxmanniaceae)
Asparagoideae (= Asparagaceae s.s.)
Nolinoideae (= Ruscaceae)
A árvore mostrada acima pode ser dividida nos seguintes arupamentos: (1) o grupo parafilético basal, as 'Asparagales inferiores' (ou 'asparagoides inferiores'); (2) o grupo das Orchidaceae às Asphodelaceae;[73] e (3) um grupo residual monofilético designado por 'Asparagales nucleares' (ou 'asparagoides superiores'), compreendendo as duas famílias maiores, Amaryllidaceae sensu lato e Asparagaceae sensu lato.[1]
As duas características diferenciadoras entre esses dois grupos (embora com excepções) são: (1) o modo de microsporogénese; e (2) a posição do ovário. As 'Asparagales inferiores' normalmente têm microsporogénese simultânea (ou seja, as paredes celulares desenvolvem-se apenas após ambas as divisões meióticas), o que parece ser uma apomorfia dentro das monocotiledóneas, enquanto os 'Asparagales centrais' reverteram para a microsporogénese sucessiva (isto é, as paredes celulares desenvolvem-se após cada divisão)[60] As 'Asparagales inferiores' normalmente apresentam ovário inferior, enquanto as 'Asparagales centrais' reverteram para ovário superior.
Um estudo morfológico de 2002, realizado pela filogenista britânica Paula Rudall, considerou que a presença de um ovário inferior era uma sinapomorfia das Asparagales, postulando que a reversão para um ovário superior nas 'Asparagales nucleares' poderia estar associada à presença de nectários abaixo dos ovários.[74] No entanto, outros investigadores observaran que os ovários superiores estão distribuídos entre os 'Asparagales inferiores' de tal forma que não é claro onde colocar a evolução das diferentes morfologias ováricas. A posição do ovário parece um carácter muito mais flexível (aqui e em outras angiospermas) do que se pensava anteriormente.[1]
O sistema APG IV, publicado em 2016, manteve em relação às monocotiledóneas em geral, e em relação às Asparagales em particular o essencial do que havia sido proposto pelo sistema APG III, de 2009, que introduzira uma grande expansão das famílias Xanthorrhoeaceae, Amaryllidaceae e Asparagaceae.[9] Em consequência dessa expansão, 13 das famílias do anterior sistema APG II foram reduzidas ao nível taxonómico de subfamílias e integradas nestas três famílias. A família Xanthorrhoeaceae expandida foi redesignada como a família Asphodelaceae, e para tornar o nome Asphodelaceae disponível para uso quando esta família em sentido estrito é combinada com Xanthorrhoeaceae, a conservação de Asphodelaceae foi proposta.[75] e aprovada pelo Comité de Nomenclatura para Plantas Vasculares.[76][77] O quadro que se segue compara as famílias como estabelecidas no sistema APG II (à esquerda) com as suas equivalentes subfamílias nos sistemas APG III e APG IV (à direita):
Orchidaceae é a maior das famílias de angiospermas e, portanto, de longe, a maior da ordem Asparagales. O sistema Dahlgren reconhecia três famílias de orquídeas, mas a análise da sequência de DNA veio posteriormente mostrou que essas famílias eram polifiléticas e, portanto, deveriam ser combinadas. Vários estudos sugerem (com alto suporte de bootstrap) que a família Orchidaceae é o grupo-irmão do resto das Asparagales.[62][63][64][78] Outros estudos colocaram as orquídeas de forma diferente na árvore filogenética, geralmente dentro do clado «Boryaceae a Hypoxidaceae».[79][60][55][80][81] Em consequência, a posição de Orchidaceae mostrada acima parece a melhor hipótese atual,[1] mas não pode ser tida por confirmada.
As orquídeas apresentam microsporogénese simultânea e ovários ínferos, duas características típicas das «Asparagales inferiores». No entanto, os seus nectários raramente estão nos septos dos ovários, e a maioria das orquídeas apresenta sementes minúsculas, parecidas com pó, atípicas do resto da ordem, embora alguns membros de Vanilloideae e Cypripedioideae tenham sementes crustosas, provavelmente associadas à dispersão por pássaros e mamíferos que são atraídos pela fermentação dos seus frutos carnudos que liberta compostos aromáticos, entre os quais a baunilha.
Em termos de número de espécies, a diversificação na família Orchidaceae é notável. No entanto, embora os outras Asparagales possam ser menos ricos em espécies, são mais variáveis morfologicamente, incluindo formas semelhantes a árvores (arborescentes).
As quatro famílias excluindo Boryaceae formam um clado bem suportado em estudos baseados na análise de sequências de DNA. Todos estas quatro famílias contêm relativamente poucas espécies, e já foi sugerido que sejam combinadas numa única família sob o nome de Hypoxidaceae sensu lato.[82] A relação entre a família Boryaceae (que inclui apenas dois géneros, Borya e Alania) e as restantes Asparagales permaneceu obscura por um longo tempo. As Boryaceae são plantas que formam associações micorrizais, mas não da mesma forma que as orquídeas. Estudos morfológicos sugeriram uma relação estreita entre as Boryaceae e as Blandfordiaceae,[60] pelo que se reconhecer existir um suporte relativamente baixo para a posição de Boryaceae na árvore filogenética mostrada acima.[62]
A relação mostrada entre Ixioliriaceae e Tecophilaeaceae ainda não está clara. Alguns estudos sustentaram a existência de um clado formado por essas duas famílias,[62] mas outros estudos não encontraram suporte suficiente para considerar válida essa relação.[79] A posição da família Doryanthaceae também variou, com suporte para a posição mostrada acima,[63] mas também existem dados que dão suporte para outras posições.[62]
O clado da família Iridaceae para cima parece ter um suporte mais forte. Todos os seus membros apresentam algumas características genéticas em comum, tendo perdido os telómeros do tipo Arabidopsis.[83] A família Iridaceae distingue-se entre os Asparagales pela sua estrutura única da inflorescência (um ripídio), pela combinação de um ovário ínfero e três estames e pela ocorrência comum de folhas unifaciais, enquanto as folhas bifaciais são a norma nas outros Asparagales.
Os membros do clado de Iridaceae para cima têm nectários septais infra-loculares, que são interpretados como um direccionador para ovários secundariamente súperos.[74]
O nó seguinte na árvore filogenéticas das Asparagales (Xanthorrhoeaceae sensu lato + 'Asparagales nucleares') tem suporte forte nos dados filogenéticos disponíveis.[84] O espessamento secundário do tipo 'anómalo' ocorre entre os membros deste clado, por exemplo no género Xanthorrhoea (família Asphodelaceae) e em Dracaena (família Asparagaceae sensu lato), com espécies arborescentes que atingem proporções semelhantes a árvores de médio porte.
O agrupamento designado por 'Asparagales nucleares' (ou 'Asparagales centrais'), englobando as Amaryllidaceae sensu lato e as Asparagaceae sensu lato, formam um clado fortemente suportado nos dados filogenéticos conhecidos,[63] tal como o são os clados que constituem cada uma das famílias na sua presente circunscrição. As relações dentro dessas famílias amplamente definidas parecem, contudo, menos claras, particularmente dentro das Asparagaceae sensu lato. Observa-se que a maioria das suas subfamílias são difíceis de reconhecer e que divisões significativamente diferentes foram usadas no passado, de modo que o uso de uma família amplamente definida para se referir a todo o clado é justificado.[1] Em consequência, as relações entre subfamílias mostradas acima, com base na APWeb são um tanto incertas.
Vários estudos tentaram datar a evolução das Asparagales com base em evidências filogenéticas. Estudos mais antigos[85][86] geralmente fornecem datas mais recentes do que estudos posteriores,[79][87] tendo estes estudos mais modernos sido preferidos na tabela abaixo.
Um estudo de 2009 sugere que as Asparagales têm a maior taxa de diversificação nas monocotiledóneas, quase igual à ordem Poales, embora em ambas as ordens a taxa seja pouco mais da metade da que ocorre em Lamiales, uma ordem de eudicotiledóneas, o clado com a taxa de diversificação mais alta que se conhece.[87]
A diversidade taxonómica das monocotiledóneas foi descrita em detalhe por Klaus Kubitzki.[88][89] Informações actualizadas sobre as Asparagales podem ser encontrado no site de filogenia de angiospermas.[1]
As circunscrições familiares do sistema APG IV são as usadas como base da Lista de Verificação Mundial de Famílias de Plantas Selecionadas (a World Checklist of Selected Plant Families) mantida pelo Jardim Botânico de Kew.[90] Com esta circunscrição, a ordem consiste em 14 famílias (o sistema Dahlgren tinha 31)[91] com aproximadamente 1120 géneros e 36 000 espécies.[1] A estrutura da ordem é a seguinte:
As versões do sistema APG anteriores a 2003 (os APG e APG II) permitiam famílias 'entre colchetes', ou seja, famílias que poderiam ser segregadas de famílias mais abrangentes ou poderiam ser incluídas nelas. Estas são as famílias indicadas em "incluindo" na lista acima. A partir da sua versão APG III, o sistema APG não permite famílias entre parênteses, exigindo o uso da família mais abrangentes. Com essa excepção, a circunscrição dos Asparagales permaneceu inalterada desde 2003. Um documento separado que acompanha a publicação do sistema APG III (de 2009) forneceu subfamílias para acomodar as famílias que foram descontinuadas.[92] O primeiro sistema APG (de 1998) continha algumas famílias extras, incluídas na lista acima com indicação da família a que foram agregadas.
O sistema de Cronquist não reconhecia a ordem Asparagales, colocando a maioria das plantas envolvidas na ordem Liliales, com alguns dos géneros incluídos na própria família Liliaceae. Os sistemas mais antigos que usam a ordem Asparagales são o sistema Dahlgren,[59] de 1985, e o sistema Kubitzki,[88] de 1998. As famílias incluídas nas circunscrições da ordem usadas nestes dois sistemas são apresentadas na primeira e segunda colunas da tabela abaixo. A família equivalente no sistema APG IV (veja abaixo) é mostrada na terceira coluna. Observe que embora esses sistemas possam usar o mesmo nome para uma família, os géneros que ele inclui podem ser diferentes, de modo que a equivalência entre os sistemas é apenas aproximada em alguns casos.
a) O sistema de Dahlgren colocava esta ordem na super-ordem Lilianae, subclasse Liliidae, classe Magnoliopsida, usando a circunscrição constante da tabela.
A família das orquídeas é uma das duas maiores famílias de angiospermas (a outra é Asteraceae). A forma das flores é muito distinta, tornando as orquídeas fáceis de reconhecer. A flor é bilateralmente simétrica. As três sépalas são geralmente coloridas e brilhantes (é por isso que às vezes são chamadas de tépalas externas), com uma de cada lado ("sépalas laterais") e uma geralmente no topo da flor ("sépala dorsal"), às vezes formando um capuz. As três pétalas (ou tépalas internas), também vistosas, estão localizadas alternadamente entre as sépalas, duas em posição lateral e uma geralmente na parte inferior da flor. A pétala inferior é conhecida como labellum ou labelo, e geralmente é distintamente diferente das pétalas laterais. O género Thelymitra é um exemplo de um táxon onde a pétala inferior é semelhante em aparência às outras pétalas. Os sistemas de polinização das orquídeas estão entre os mais complexos e interessantes de todas as angiospermas. As orquídeas incluem muitas espécies de grande valor ornamental. A baunilha é obtida do fruto da orquídea Vanilla planifolia.
O género Borya contém espécies arborescentes, semelhantes a árvores de médio porte. Os membros desta famílias comportam-se como «plantas de ressurreição», pois crescendo em encostas rochosas e expostas ao sol, as plantas secam durante a estação seca e adquirem uma cor laranja enferrujada, mas tornam-se rapidamente verdes e ativas quando começa a chover. Junto com o outro género da família Boryaceae, o género Alania, estas plantas xerófitas são nativas da Austrália.
Blandfordia é o único género na família monotípica Blandfordiaceae, com quatro espécies distribuídas pelo leste da Austrália. São comummente chamados de "sinos-de-natal", devido ao formato de suas flores e à época de floração, que coincide com o Natal na Austrália. São ervas perenes, verticais, com até cerca de 1,50 m de altura, com folhas distintas. A inflorescência é um racemo. As flores individuais têm pedicelos e tépalas articulados, formando uma estrutura tubular. As sementes apresentam tricomas conspícuos.
Lanaria lanata é a única espécie da família monotípica Lanariaceae. A espécie tem distribuição natural no sul da África do Sul. Sendo uma monocotiledónea mais ou menos típica, a espécie pode ser reconhecida pela sua inflorescência pouco ramificada coberta por pêlos ramificados (dando origem ao nome comum de cauda de cordeiro). As flores são radialmente simétricas.
Asteliaceae é uma família com dois a quatro géneros, com distribuição natural no hemisfério sul. São espécies mais ou menos rizomatosas, com folhas que seguem uma filotaxia em espiral. Produzem uma inflorescência que pode formar um racemo ou espigo. Apresenta grandes brácteas na base da inflorescência. As flores individuais são pequenas, com tépalas unidas na base.
A família inclui cerca de 150 espécies com distribuição cosmopolita, excluindo Europa e norte da Ásia. As espécies podem ser reconhecidas pela sua roseta de folhas mais ou menos dobradas, com bases persistentes e pêlos não-glandulares bastante proeminentes. As tépalas no verticilo externo tendem a ser verdes por fora. O ovário é ínfero, muitas vezes com uma porção tubular delgada no seu ápice formada por tépalas unidas ou na ponta do ovário.
A família inclui um único género, Ixiolirion, com quatro espécies distribuídas do Egipto à Ásia Central. Estas espécies são plantas herbáceas com cormos, que produzem uma inflorescência em cacho. As flores individuais são azuis, tubulares na base, com ovário ínfero.
Os nove géneros que integram a família Tecophilaeaceae são encontrados no Chile, nos Estados Unidos e na África. São ervas com cormos e com folhas que às vezes são pendentes (pecioladas) e com lâminas largas. As flores têm tépalas que se abrem para fora. Os estames são fortemente dimórficos. A antera é aberta por poros. O género Cyanastrum às vezes é colocado numa família autónoma designada por Cyanastraceae.
As duas espécies de Doryanthes, o único género da família, são ervas de grande porte, formadoras de rosetas, que são um elemento conspícuo da flora nas proximidades de Sydney, Austrália, sendo difícil não as notar quando em flor. As folhas têm margens inteiras, mas se desintegram-se em fibras no ápice. As inflorescências sub-umbeladas inserem-se no ápice de hastes longas, contendo numerosas flores vermelhas brilhantes, que são radialmente simétricas com ovários ínferos.
A família Iridaceae contém cerca de 70 géneros e mais de 1 600 espécies, com distribuição cosmopolita. Os membros da família são geralmente ervas perenes, com folhas unifaciais em forma de espada (ensiformes). A inflorescência é uma espiga ou panícula de flores solitárias, ou forma um monocásio cimoso ou um ripídio (significando que as hastes sucessivas das flores seguem um caminho em ziguezague no mesmo plano). A flor tem apenas três estames, cada um oposto a uma tépala externa.
O açafrão é obtido a partir dos estilos secos de Crocus sativus L., um membro da família da íris. Os cormos de algumas espécies de Iridaceae são usados como alimento por alguns povos indígenas. Muitas espécies da família da íris têm grande importância económica na horticultura ornamental e na indústria de flores de corte, especialmente Gladiolus, Freesia, Sparaxis , Iris, Tigridia (lírio-tigre), Ixia (lírio-do-milho), Romulea, Neomarica, Moraea (lírio-borboleta), Nemastylis, Belamcanda, Sisyrinchium (grama-de-olhos-azuis), Crocosmia e Trimezia. Muitos outros géneros, tanto perenes como bolbosos, são cultivados em jardins nas regiões tropicais e temperadas (por exemplo, Watsonia, Crocus, Dietes, Tritonia, Hesperantha e Neomarica).[93][94][95] Os géneros Moraea e Homeriasão plantas venenosas que são um problema nas regiões produtoras de ovelhas e gado bovino, principalmente na África do Sul.[93]
A família consiste num único género Xeronema com duas espécies, uma encontrada apenas nas ilhas Poor Knights da Nova Zelândia e a outra na Nova Caledónia. A inflorescência é repleta de flores bastante grandes, radialmente simétricas, voltadas para cima. Os estames são fortemente exsertos (ou seja, estendem-se para fora da flor). A família é pouco conhecida.
As Asphodelaceae, ou família do asfodelo, foi reconhecida, embora de forma e com circunscrição variadas, por muitos taxonomistas, mas os limites da família variaram muito. Com base nos resultados obtidos pelas técnicas da filogenética, o sistema APG III, de 2009, da classificação do APG, agrupou as antigas famílias Hemerocallidaceae, Xanthorrhoeaceae sensu stricto e Asphodelaceae sensu stricto numa família mais ampla a que foi dada a designação de Xanthorrhoeaceae.[58] Um artigo publicado na mesma época propôs que as três famílias originais deveriam ser mantidas como subfamílias dentro de Xanthorrhoeaceae sensu lato.[92] O nome «Xanthorrhoeaceae» tem prioridade como nome de família, mas foi considerada como não refletindo a natureza da família, e em 2014 foi acordado propor ao Congresso Internacional de Botânica de 2017 a conservação formal do nome Asphodelaceae em detrimento de «Xanthorrhoeaceae», o que foi aprovado por aquele órgão, que é responsável pela nomenclatura vegetal. O sistema APG IV usa o nome Asphodelaceae.[96]
A família da amarílis foi reconhecida em muitos sistemas taxonómicos, mas os limites da família variaram ao longo dos tempos. Na definição mais restrita, Amaryllidaceae sensu stricto é caracterizada por uma inflorescência umbelada com um ovário ínfero. Dois outros grupos têm inflorescências semelhantes, mas um ovário súpero, pelo que às vezes foram colocados em famílias separadas: as famílias Agapanthaceae e Alliaceae. Com base na pesquisa filogenética, a revisão mais recente (2016) da classificação APG agrupa essas três famílias sob o nome conservado de Amaryllidaceae.[58] Antes da sua revisão de 2009, o APG havia usado o nome Alliaceae para este grupo.[54] Um artigo publicado ao mesmo tempo que a classificação de 2009 propôs que as três famílias originais deveriam ser mantidas como subfamílias dentro de Amaryllidaceae sensu lato,[92] o que está consagrado no sistema APG IV e é aqui usado.
Os membros deste agrupamento têm uma história taxonómica complexa, tendo sido atribuídos a famílias amplamente diferentes em diferentes sistemas de classificação. Os subgrupos propostos são difíceis de reconhecer, tendo flores semelhantes a lírios, com o resultado de que alguns membros do grupo foram incluídos em subgrupos diferentes em momentos diferentes.[98] Com base em estudos de filogenética, a última revisão (2016) da classificação APG suporta o uso de uma única família amplamente definida, Asparagaceae sensu lato.[58] Um artigo publicado ao mesmo tempo que a classificação de 2009 propôs sete subfamílias para as famílias reconhecidas na primeira classificação APG de 1998.[92] Esta divisão foi mantida no sistema APG IV e foi usado aqui, embora não esteja claro se a abordagem será confirmada por investigação futura, já que alguns dos clados têm suporte insuficiente. A família definida de forma alargada é grande, com cerca de 153 géneros e mais de 2480 espécies, e ocorre em todo o mundo. As sete subfamílias parecem formar dois grupos: (1) [Aphyllanthoideae + Brodiaeoideae + Scilloideae + Agavoideae], com poucas características morfológicas comuns que as diferenciem da família como um todo; e (2) [Lomandroideae + Asparagoideae + Nolinoideae], com algumas características morfológicas comuns.[98][99]
A ordem Asparagales inclui muitas plantas de cultivo e plantas ornamentais importantes. As culturas incluem Allium (alhos e cebolas), Asparagus (espargos) e Vanilla (baunilha), enquanto as plantas ornamentais incluem os íris, jacintos e orquídeas.[10]
Aspargales é uma ordem de plantas com flor que na circunscrição taxonómica que lhe é dada pelo sistema de classificação APG IV (2016) é a maior das ordens de monocotiledóneas, agrupando 14 famílias, com 1122 géneros e cerca de 36 000 espécies, entre as quais algumas com elevado interesse económico, como os espargos, as cebolas, o sisal, a baunilha e múltiplas plantas ornamentais. Apesar de claramente circunscrita com base na sua filogenia molecular, a ordem é difícil de definir morfologicamente, pois os seus membros são estruturalmente muito diversos, indo dos pequenos caméfitos às lianas e a alguns arbustos e pequenas árvores, embora contenha maioritariamente espécies herbáceas perenes com elevada proporção de geófitos.
Sparrisordningen (Asparagales) är en ordning av enhjärtbladiga växter som omfattar ett antal familjer med växter som inte är träd. I äldre klassificeringssystem ingick dessa familjer i ordningen Liliales och några släkten ingick till och med i liljeväxtfamiljen. I vissa system utgör Orchidales och Iridales egna ordningar, men speciellt klassificeringssystemet Angiosperm Phylogeny Group inkluderar dessa i Asparagales.
Asparagales har fått sitt namn efter släktet sparrisar som har det vetenskapliga namnet Asparagus.
Sparrisordningen (Asparagales) är en ordning av enhjärtbladiga växter som omfattar ett antal familjer med växter som inte är träd. I äldre klassificeringssystem ingick dessa familjer i ordningen Liliales och några släkten ingick till och med i liljeväxtfamiljen. I vissa system utgör Orchidales och Iridales egna ordningar, men speciellt klassificeringssystemet Angiosperm Phylogeny Group inkluderar dessa i Asparagales.
Asparagales har fått sitt namn efter släktet sparrisar som har det vetenskapliga namnet Asparagus.
Холодкоцвіті (Asparagales) — порядок квіткових рослин класу монокотів.
Серед представників ряду кілька їстивних рослин, зокрема родів цибуля і спаржа. Багато представників вирощуються як декоративні, завдяки своїм квіткам або як кімнатні рослини: конвалія, проліска, нарцис, амариліс, орхідеї, агава, алое, юка.
Холодкоцвіті (Asparagales) — порядок квіткових рослин класу монокотів.
Серед представників ряду кілька їстивних рослин, зокрема родів цибуля і спаржа. Багато представників вирощуються як декоративні, завдяки своїм квіткам або як кімнатні рослини: конвалія, проліска, нарцис, амариліс, орхідеї, агава, алое, юка.
Bộ Măng tây hay bộ Thiên môn đông (danh pháp khoa học: Asparagales) là một bộ trong lớp thực vật một lá mầm bao gồm một số họ cây không thân gỗ. Trong các hệ thống phân loại cũ, các họ mà hiện nay đưa vào trong bộ Asparagales đã từng được đưa vào trong bộ Loa kèn (Liliales), và một số chi trong đó thậm chí còn được đưa vào trong họ Loa kèn (Liliaceae). Một số hệ thống phân loại còn tách một số họ được liệt kê dưới đây thành các bộ khác, bao gồm cả các bộ Orchidales (lan) và bộ Iridales (diên vĩ), trong khi các hệ thống khác, đặc biệt là hệ thống phân loại của APG lại đưa hai bộ Orchidales và Iridales vào trong bộ Asparagales. Bộ này được đặt tên theo chi Asparagus (măng tây, thiên môn đông).
Hệ thống APG III năm 2009 công nhận các họ sau:
nghĩa là hai họ Alliaceae và Agapanthaceae nhập vào trong họ Amaryllidaceae và không còn có các họ tùy chọn tách ra từ hai họ Asparagaceae và Xanthorrhoeaceae nữa.
Hệ thống phân loại của Angiosperm Phylogeny Group được các nhà thực vật học sử dụng rộng rãi và đã được cập nhật thành APG II năm 2003 để thêm vào các nghiên cứu gần đây, đặc biệt là các nghiên cứu phân tích ADN. Sơ đồ năm 1998 xác định 29 họ trong bộ Asparagales. APG II hợp nhất một số họ và thừa nhận một hệ thống khác với ít số lượng họ nhưng họ lại lớn hơn, trong đó một số họ nhỏ có thể kết hợp lại thành các họ lớn hơn, dựa trên sự tương thích di truyền gần gũi hơn và vẫn tuân thủ 'hệ thống APG', cho rằng họ này chứa 16-24 họ với 1.122 chi và khoảng 26.070 loài. Ví dụ, theo hệ thống phân loại mới hơn này thì người ta có thể đưa chính xác cây hoa hiên hay cây kim châm (chi Hemerocallis) vào họ Hemerocallidaceae, hoặc vào họ Xanthorrhoeaceae. Phân loại của APG II về bộ Asparagales như sau:
Hệ thống APG năm 1998 cũng đặt bộ này trong nhánh thực vật một lá mầm nhưng sử dụng định nghĩa sau:
Các hệ thống phân loại tách ba bộ Măng tây (Asparagales), bộ Phong lan (Orchidales) và bộ Diên vĩ (Iridales) nói chung được sắp xếp như sau:
Hệ thống Dahlgren đặt bộ này trong siêu bộ Lilianae thuộc phân lớp Liliidae [= thực vật một lá mầm] của lớp Magnoliopsida [= thực vật hạt kín] và sử dụng định nghĩa sau:
Hệ thống Cronquist không công nhận bộ này, đặt nhiều chi trong bộ Liliales (phân lớp Liliidae của lớp Liliopsida [= thực vật một lá mầm]). Một số chi được đặt trong họ Liliaceae.
Hệ thống Wettstein, phiên bản cuối cùng năm 1935, cũng không công nhận bộ này và đặt nhiều chi trong bộ Liliiflorae thuộc lớp Monocotyledones.
Cây phát sinh chủng loài của bộ Măng tây so với các bộ thực vật một lá mầm khác lấy theo APG III.
Monocots
Asparagales
Commelinidae (commelinids)
Cây phát sinh chủng loài trong nội bộ bộ Măng tây lấy theo APG III.
Asparagales
Xanthorrhoeaceae s.l.
Hemerocallidoideae (Hemerocallidaceae)
Xanthorrhoeoideae (Xanthorrhoeaceae s. s.)
Asphodeloideae (Asphodelaceae)
Amaryllidaceae s.l.
Agapanthoideae (Agapanthaceae)
Amaryllidoideae (Amaryllidaceae s. s.)
Asparagaceae s.l.
Aphyllanthoideae (Aphyllanthaceae)
Asparagoideae (Asparagaceae s. s.)
Bộ Măng tây hay bộ Thiên môn đông (danh pháp khoa học: Asparagales) là một bộ trong lớp thực vật một lá mầm bao gồm một số họ cây không thân gỗ. Trong các hệ thống phân loại cũ, các họ mà hiện nay đưa vào trong bộ Asparagales đã từng được đưa vào trong bộ Loa kèn (Liliales), và một số chi trong đó thậm chí còn được đưa vào trong họ Loa kèn (Liliaceae). Một số hệ thống phân loại còn tách một số họ được liệt kê dưới đây thành các bộ khác, bao gồm cả các bộ Orchidales (lan) và bộ Iridales (diên vĩ), trong khi các hệ thống khác, đặc biệt là hệ thống phân loại của APG lại đưa hai bộ Orchidales và Iridales vào trong bộ Asparagales. Bộ này được đặt tên theo chi Asparagus (măng tây, thiên môn đông).
Спаржецветные (лат. Asparagales) — порядок однодольных растений. В системе классификации APG II включён в группу монокоты.
Наиболее известные пищевые растения из этого порядка — представители родов лук и спаржа.
Среди спаржецветных много декоративных красивоцветущих растений, например, ландыш, нарцисс, амариллис, орхидеи и другие.
Комнатные растения, входящие в этот порядок: агава, алоэ, юкка.
Согласно Системе APG II (2003) порядок Спаржецветные (Asparagales) состоит из 25 семейств:
В системе классификации Кронквиста (1981) порядок Asparagales отсутствует. Большинство таксонов, включаемых в этот порядок в Системе APG II, в Системе Кронквиста входит в порядок Liliales подкласса Liliidae класса Liliopsida.
В системе классификации Тахтаджяна (1997) порядок Спаржецветные (Asparagales) входит в состав надпорядка Lilianae подкласса Liliidae класса Liliopsida.
Спаржецветные (лат. Asparagales) — порядок однодольных растений. В системе классификации APG II включён в группу монокоты.
天門冬目(学名:Asparagales)是被子植物中的單子葉植物。傳統上本目的植物常被分在廣義百合目或蘭目中,現代的植物分類學者多根据《被子植物APG分类法》(1998年)、《被子植物APG II分类法 (修订版)》(2003年)、《被子植物APG III分类法》(2009年) 採用1829年林克所發表的學名。
依據《被子植物APG III分类法》(2009年)[1],天門冬目有下列各科::
鸭跖草目 Commelinales
薑目 Zingiberales
禾本目 Poales
棕榈目 Arecales
天门冬目 Asparagales
百合目 Liliales
薯蓣目 Dioscoreales
露兜树目 Pandanales
无叶莲目 Petrosaviales
澤瀉目 Alismatales
菖蒲目 Acorales
天門冬目(学名:Asparagales)是被子植物中的單子葉植物。傳統上本目的植物常被分在廣義百合目或蘭目中,現代的植物分類學者多根据《被子植物APG分类法》(1998年)、《被子植物APG II分类法 (修订版)》(2003年)、《被子植物APG III分类法》(2009年) 採用1829年林克所發表的學名。
 分類 界 : 植物界 Plantae 階級なし : 被子植物 Angiosperm 階級なし : 単子葉類 Monocots 目 : キジカクシ目 Asparagales 学名 Asparagales Diels (1917) 和名 キジカクシ目[1][2]、クサスギカズラ目[2]
分類 界 : 植物界 Plantae 階級なし : 被子植物 Angiosperm 階級なし : 単子葉類 Monocots 目 : キジカクシ目 Asparagales 学名 Asparagales Diels (1917) 和名 キジカクシ目[1][2]、クサスギカズラ目[2] 
キジカクシ目(キジカクシもく、学名:Asparagales)は、単子葉植物の目の1つ。クサスギカズラ目[2]とも。
APG体系では、ラン科やアヤメ科など、単子葉植物の14科を含む大きな目である[3]。科の和名は、大場(2009)に基づく[1]。
クロンキスト体系ではキジカクシ科を独立の科としておらず、キジカクシ目も存在しない。APGでキジカクシ目に属している多くの科はユリ目(多くはユリ科)に入れられている。
単子葉植物が専門であったダールグレン体系ではキジカクシ目を置き、科レベルでも細分している。(ただしアヤメ科やラン科はここに入れず、各科の範囲もAPGとは異なる)
新エングラー体系ではキジカクシ科を独立の科としておらず、キジカクシ目も存在しない。APGでキジカクシ目に属している多くの科はユリ目(多くはユリ科)に入れられている。
キジカクシ目(キジカクシもく、学名:Asparagales)は、単子葉植物の目の1つ。クサスギカズラ目とも。
비짜루목 또는 아스파라거스목(Asparagales)은 외떡잎식물군에 속하는 속씨식물 목이다. 이 목에 비짜루과(아스파라거스과)는 반드시 포함되나, 다른 과들은 분류 체계에 따라 다르게 포함되기도 한다. 확실한 것은 아니지만 백악기 사이에 갈라져 나와 진화한 것으로 추측하고 있다. 그러나 과들의 상이한 분류 체계때문에 이들의 진화는 전적으로 확실한 것은 아니다.
2003년의 APG II 분류 체계에서, 비짜루목은 아래와 같이 외떡잎식물군으로 분류된다.:
참고: "+ ..." = 앞에 기술된 과에서 분리하기도 하는, 선택적으로 분리되는 과.
다음은 외떡잎식물군의 계통 분류이다.[1]
외떡잎식물군 닭의장풀군1998년의 APG 분류 체계도 또한 아래와 같이 외떡잎식물군으로 분류하고, 상세 분류는 아래와 같다:
1998년판의 쿠비츠키 분류 체계에서는 아래와 같이 분류하였다.
달그렌 식물 분류 체계에서는 백합아강(Liliidae)의 백합아목(Lilianae)으로 분류하였고, 상세한 과 분류는 아래와 같다:
크론퀴스트 식물 분류 체계는 목으로 간주하지 않고, 대부분을 (백합아강 하위의) 백합목에 포함시킨다. 일부의 속은 백합과에 포함되기도 한다. 1935년 최종판의 베트슈타인 식물 분류 체계에서는 하나의 목으로 인정하지 않고, 대부분의 식물 종들을 외떡잎식물강의 백합목(Liliiflorae)으로 분류하였다.


















_-_8.jpg)







_12_ies.jpg)































_-_8.jpg)





_12_ies.jpg)




































_12_ies.jpg)
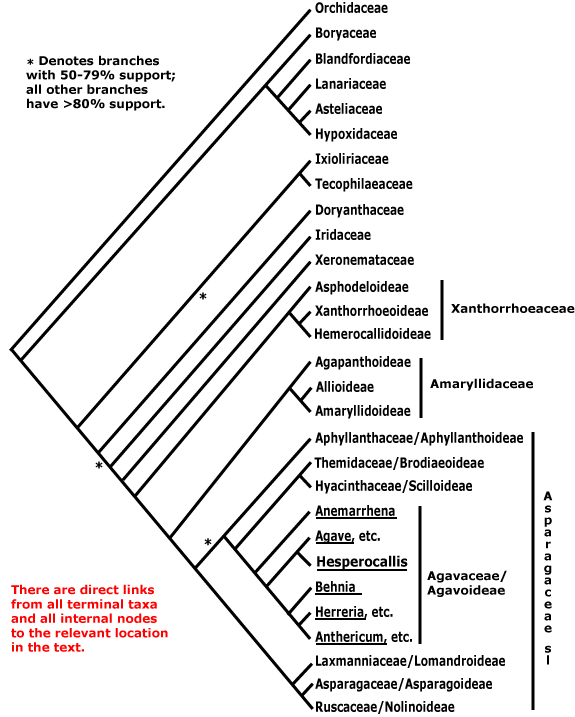{kind=link}