“Batoteuthis skolops, new species
External Characteristics
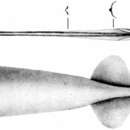
The Holotype. The mantle wall is very thin but muscular, similar to the mantle wall of most cranchiids. The mantle is sac-like and bulges in the middle, but it tapers to a point posteriorly; its muscular portion terminates just anterior to the posterior margin of the fins. The free mantle opening is very broad and is considerably wider than the head. The gladius is visible in the dorsal mid-line and extends posteriorly beyond the fins as the supporting element of the long, thin "tail", much of which is missing (Pl. I, Figs. A, B).
The fins are small and short; their length is about half their width. Taken together, the 2 fins are broadly oval in outline. Anterior lobes are not developed, and the posterior margins extend for a short distance along the narrow tail.
Because of the peculiar long, tapering tail-like posterior extensions of the mantle and fins, it is necessary to measure mantle and fin length exclusive of this "tail"; therefore the mantle and fin length are defined
TABLE 1. List of Material of Batoteuthis skolops, New Species
Eltanin Sta. No.a
Mantle Length, mm
Sex
Location
Apparent Depth of Capture, m
Date of Capture
Gear
1396b
51
Female?
49º57’S, 169º01’W
2525
26 XI 1964
3m IKMT
1314
30
?
59º11’S, 105º04’W
366
24 X 1964
3m IKMT
846
95
Female
57º52’S, 74º43’W
1866
10-11 XI 1963
3m IKMT
382
90
Female
55º23’S, 55º45’W
1257
23 XII 1962
3m IKMT
a These represent the University of Southern California’s station numbers assigned for Eltanin cruises.
b Holotype.
here as extending to the level of greatest inflection on the concave posterior border of the fins. It must be emphasized that the method of measurement differs from the usual method in which the tail, if present, is included in the mantle length. Whenever the term mantle length is used here with reference to the Batoteuthidae, it refers to the convenient definition stated above.
The funnel is huge and thin walled like the mantle; its exhalant opening is very broad. The funnel extends nearly to the base of the ventral arms (Pl. I, Fig. A). The dorsal member of the funnel organ is an inverted U-shaped structure that has an inverted V-shaped inner notch. A small papilla extends anteriorly from the apex. The ventral members are basically oval but exhibit some variation in shape (Pl. IV, Fig. G). The funnel valve is the typical semicircular flap.
The funnel component of the locking apparatus is nearly straight, deeply grooved, and slightly broader and deeper in the posterior end. The mantle component extends posteriorly from the free edge of the mantle as a nearly straight, narrow ridge which broadens posteriorly (Pl. IV, Fig. D).
The head is exceedingly small and is slightly shorter than broad. The eyes are proportionately large and occupy about ¾ of the lateral surface of the head. The eye opening is oval with a small anterior optic sinus. Small olfactory papillae are present just posterior to the eye on the ventrolateral surface of the head. Both nuchal crest and nuchal folds are absent.
Arms are long and slender they taper distally to terminate in long, attenuate tips. The arm formula is 4.3.2.1. No evidence of swimming keels exists except on the ventral arms, where keels are weakly developed. Trabeculae and sucker stalks arise from a common base (Pl. II, Fig. D). The protective membranes are variable but generally very low and weakly developed. However, the membranes appear to be more strongly developed on the right ventral arm of the type specimen, where they reach as high as the trabeculae.
Suckers are arranged in well separated, biserial rows and stand considerably above the oral surface of the arms on long stalks. The slender portion of the sucker stalk arises abruptly from a broad, conical base. At the tips of the arms the size and arrangement of the suckers cannot be determined because of damage to the specimen.
The largest suckers of the second and third arms are approximately ½ greater in diameter than the largest suckers of the first and fourth arms. The globular arm suckers possess broad apertures. The rings of the lateral arm suckers possess approximately 8-10 large, broadly spaced, sharply pointed, conical teeth, while the sucker rings of the dorsal and ventral arms possess 11-12 small, closely packed, sharp teeth (Pl. II, Figs. B, F, G, H, I) .
Tentacles are relatively short (about 1 ½ times longer than the arms) and robust. The extremely long clubs occupy approximately 80 % of the tentacle length; they are flattened but not appreciably expanded (Pl. I, Fig. C; Pl. II, Fig. E). Numerous pointed trabeculae are distributed along the entire lateral margins of the club. A trace of a protective membrane can be detected between the trabeculae, especially between the first few proximal trabeculae on the ventral margin of the club. The clubs have 6 well defined longitudinal rows of small, closely packed suckers that are situated on long slender stalks. The chitinous rings bear approximately 6 small, conical, widely spaced teeth (Pl. II, Fig. A). The bases of the stalks of the marginal sucker rows join with the bases of the trabeculae (Pl. II, Fig. C).
No dactyl or carpal areas are present on the club. The 2 median sucker rows and the lateral-most rows appear to originate in the same proximal region of the club, while the remaining 2 rows seem to converge proximally with the lateral-most rows.
TABLE 2. Measurements and Indicesa of Batoteuthis skolops, New Species
Elt. 1396 Holotype Female?
Elt. 1314 Paratype ?
Elt. 846 Female
Elt. 382 Female
Measure-ments, mm
Indices
Measure-ments, mm
Indices
Measure-ments, mm
Indices
Measure-ments, mm
Indices
Mantle Lengthb
51
…
30
…
95
…
90
…
Tail Length
10+c
…
25.5+
85.0
42+
…
29+
…
Mantle Width Opening
14
27.4
7
23.4
13
13.7
15
16.7
Mantle Width Widest
19
37.2
12
40.0
20
21
18
20
Head Width
8.5
16.6
4.5
15.0
Missing
…
Missing
…
Fin Length
9
18.0
4
13.3
20
21
18
20
Fin Width
18
35.2
7
23.4
40
42.1
33
36.7
R.
L.
R.
L.
R.
L.
R.
L.
Arms
Id
14
13+
27.9
25.5
4.5
3.5+
15.0
11.7
Missing
…
Missing
…
II
18
17+
35.2
33.4
4+
4.5+
13.3
15.0
“
…
“
…
III
19.5
20
38.2
39.2
5.5+
5.5
18.3
18.3
“
…
“
…
IV
21.5
22.5
42.1
44.2
5.5+
5.5
23.4
23.4
“
…
“
…
Tentacle Length
30
30
58.9
58.9
21
22
70.0
73.4
“
…
“
…
Club Lengthe
24
24.5
47.0
48.0
19
20
63.4
66.7
“
…
“
…
a Indices are expressed as % of mantle length.
b Mantle measured to a level with the points of greatest inflection on the concave posterior border of the fins (see text).
c A + mark indicates that a portion of the measured character is missing (e.g., arm tips) and that the complete specimen would have greater dimensions.
d Arms measured from the dorsal notch between the bases of the arms, e.g. II is measured from the notch between II and I.
e In Elt. 1314 this refers to the entire sucker-bearing portion of the tentacle (see text).
The buccal membrane possesses 7 lappets and 8 connectives which are attached dorsally to arm pairs I and II and ventrally to arm pairs III and IV. The oral surface of the buccal membrane is nearly smooth, marked only by minute crenulations (Pl. IV, Fig. A).
The beaks are figured in Pl. IV, Fig. C. Their rostra are strong and darkly pigmented, while their lamellae are extremely fragile and transparent. A definite ridge is present on the insertion plate of the gular lamella of the lower mandible.
The radula is extremely small and the finer details are difficult to detect with certainty (Pl. IV, Fig. B). There are 7 teeth in a transverse row. The first and second laterals are very small, while the third laterals are extremely elongate.
The gladius (taken from Elt. 846) is long and slender, with the anterior half consisting of only a narrow rhachis. A narrow vane arises at about the midpoint and very shortly folds ventrally with the edges fusing in the midline to form a long, hollow, tapering conus (Pl. III, Fig. B).
A few dark brown chromatophores are present on the posterior ventral surface of the mantle, around the head, along the tentacle, and particularly on the aboral surface of the club, where they are regularly spaced between the trabeculae.
Table 2 presents the measurements and indices of the specimens of Batoteuthis skolops.
Large Specimens. The shape of the mantle in the larger specimens (Elt. 846 and 382) differs greatly from that seen in the 2 younger specimens (Pl. III, Fig. A). The mantle is very elongate, slender and conical; the widest point is just posterior to the free mantle opening, which is slightly constricted. The mantle wall remains thin, though proportionately thicker than in the smaller specimens. The gladius is visible in the dorsal midline only in the anterior region and along the tail. The tail is extremely long (nearly half the mantle length) and slender, and, although the extreme tip is missing, it undoubtedly tapers to a sharp point.
The fins are short and occupy about 1/5 of the mantle length. Their posterior borders are concave and extend posteriorly to form part of the tapering tail.
The funnel is strongly developed. The locking apparatuses are similar to those of the smaller specimens. The dorsal member of the funnel organ is the same shape as in the holotype, but each ventral member has a skewed diamond shape with rounded corners (Pl. IV, Fig. H).
The head is missing from both large specimens.
The outer integument has been rubbed off from nearly the entire mantle. However, some shreds remain on the posterior mantle and under the fins to reveal numerous typical reddish-brown chromatophores.
Larval specimen. The mantle is sac-like, membranous, and transparent (Pl. III, Fig. C). The flaring mantle opening is considerably wider than the head. The gladius is clearly visible along the midline. The muscular portion of the mantle terminates shortly before the posterior border of the fins, but the gladius continues posteriorly, covered by the mantle integument, to form an extremely long, needle-like tail. The tail is nearly equal to the mantle in length. However, the tip of the pen is missing, and it is possible that the tail would be longer in uninjured specimens.
The fins are extremely small and occupy only 13 % of the mantle length. They are nearly 1 ¾ times broader than long. The fin margins are broadly ovoid and exhibit no free lobes.
The thin-walled funnel is large and extends to the level of the anterior border of the eyes. The inverted U-shaped dorsal member of the funnel organ is broad and possesses a long, flattened papilla that extends anteriorly from the apex. The ventral pads are only slightly longer than broad and appear nearly square, although the corners are rounded; there is a slight anterior bulge.
The head is extremely small and rounded. The eyes are proportionately much smaller than in the type, and they appear to be directed slightly forward.
The arms are lacking in details and it is not possible to determine the extent of development of keels and protective membranes. The suckers are situated in 2 biserial rows. The arm tips are missing. The buccal mass, surrounded by an indistinct buccal membrane, protrudes slightly from the base of the oral crown.
The tentacles are proportionately longer than in the adult and are more robust than the arms. The proportion of tentacle length to mantle length is 75 % in the larva and 59 % in the adult. The sucker-bearing portion of the larval tentacle occupies about 90 % of the tentacle length. However, only the distal 55 % of this portion bears a club structure similar to that in the larger specimen. Proximal to the distal area there is a gradual decrease in the number of sucker rows. The larval clubs have more distinct protective membranes than do those of adults.
A few scattered chromatophores are distributed over the dorsal surface of the head.
Internal Anatomy
Although the specimens were not completely dissected, it has been possible to determine the general orientation and form of parts of the major organ systems (Pl. V, Figs. A–E; Pl. VI, Figs. A–D)
Kidney (Pl. V, Fig. B). Numerous venous appendages are present along the vena cavae, as these vessels lead from the cephalic vein to the branchial hearts. Appendages also occur along the initial part of each posterior mantle vein. Two additional kidney lobes arise near the junction of the cephalic vein and the vena cavae. These peculiar lobes extend anteriorly and lie parallel to and dorsal to the intestine. Both appear to be blind extensions of the kidney. Nephridial pores are located anterior to the branchial hearts. Typical nephridial papillae are lacking, and the pores open nearly flush with the surface of the body wall ventral to the kidneys.
Branchial heart complex (Pl. V, Fig. B). The vena cavae, as they emerge from the kidney, enter the branchial hearts near their extreme anterior ends. Immediately lateral to these points the afferent branchial vessels arise from the branchial hearts and pass into the gills. A distinct swelling that appears to be a sphincter muscle surrounds each of these branchial vessels a short distance from the branchial heart. The branchial hearts are unusually elongate, oval structures with their long axes paralleling the body axis. The branchial hearts are in a peculiar position—they lie near the body midline, almost completely enclosed in the visceral-pericardial coelom, and are far removed from the posterior end of the gills. The accessory branchial hearts are attached to the posterior tips of the branchial hearts.
Gills (Pl. V, Figs. A, C). Each gill is slender and possesses approximately 40 pairs of filaments in the larger specimens. The gills are long relative to the size of the viscera; however, they are very small in comparison to the size of the body. The branchial glands are well developed. A cross section through the gill reveals a very short, blunt inner demibranch supported by a slender stylet and a long, outer demibranch which is almost completely fused to the base of the gill. The branchial canal is large.
Female reproductive system (Pl. V, Figs. A, B; Pl. VI, Figs. B, C). The visceral-pericardial coelom forms an extremely long, slender sac that extends approximately from the base of the gills posteriorly into the tail. The immature ovary is a single organ situated far posteriorly in the visceral-pericardial coelom near the anterior edge of the fins. It is boomerang-shaped and is suspended from the dorsal wall of the coelom by a short mesentery.
An oviduct lies along each anterior lateral side of the visceral-pericardial coelom adjacent to the branchial hearts. Each oviduct is subdivided into 3 distinct sections. The posterior portion arises from the coelom by a small crenulate pore just dorsal to the nidamental glands. It extends forward to the middle area as a tightly coiled duct lined by closely packed lamellae that run parallel to the duct. The funnel-shaped middle portion consists of a central duct with an outer surface of thin lamellae. These lamellae are independent of those that run along the inner surface of the first section of the oviduct. The orifice of the middle portion opens into the third, anteriormost section, which is a bilobed, elongate, hood-like structure. The inner surface of each side of the hood bears separate rows of lamellae. The hood has a broad, slit-like orifice which opens into the mantle cavity just anterior to the afferent branchial vessel.
The nidamental glands are small, narrow, cigar-shaped structures that lie near the midline ventral to the visceral-pericardial coelom and immediately posterior to the branchial hearts. These glands open into the mantle cavity via broad slit-like pores across their anterior ends.
Visceral blood vessels (Pl. V, Figs. A, B). The cephalic artery arises from the anterior end of the large heart and passes laterally around the side of the liver instead of passing directly dorsal to this organ, as is often the case in other teuthoids. The efferent branchial vessels enter the ventricle well posteriorly.
The posterior mantle artery arises from the posterior end of the ventricle and runs along the ventral surface of body wall just below the visceral-pericardial coelom. Near the level of the anterior border of the fins it sends a branch, the median mantle artery, into the ventral portion of the mantle. This very short branch runs along the anterior edge of a median membrane. This membrane is extremely narrow and barely visible. Further posteriorly the mantle artery forks to form the paired lateral mantle arteries.
The posterior mantle veins leave the kidney, pass dorsal to the nidamental glands, and run along the lateral edge of the visceral-pericardial coelom to the region of the fins, where they enter the mantle wall and the fins.
Digestive tract (Pl. V, Fig. D). The esophagus extends from the head along the dorsal surface of the liver and passes laterally around the liver near its posterior end. The esophagus enters the stomach near the midline ventral to the liver. Both stomach and caecum have been severely damaged during capture and it is not possible to give a meaningful description. The intestine leaves the stomach-caecum complex and passes almost directly forward to the anus, which lies at the base of the funnel. Anal flaps are present.
The ink sac lies embedded in the ventral surface of the liver and gives off a short duct that leads into the rectum.
The liver is very large in relation to the size of the other viscera, and its long axis coincides with the body axis.
The pancreas is composed of 2 compact and separate organs, each of which lies on the posterior ventral surface of the liver. The organs are well defined, sausage-shaped structures, and each bears a small lobe on its medial side from which the hepatic ducts probably arise, although they cannot be observed in the present material.
Mantle and fin nerves (Pl. V. Fig. E). The pallial nerve enters the stellate ganglion without giving off a distinct fin nerve, although in some specimens a few fibers may pass over the surface of the ganglion to rejoin the large nerve trunk that emerges from the posterior end of the ganglion. In the specimens where they exist, these superficial fibers probably represent at least a portion of the fin nerve fibers; the remaining fin nerve fibers pass through the ganglion. A few large nerves emerge laterally from the ganglion and pass into the anterior portion of the mantle. A large nerve trunk that incorporates both posterior mantle and fin nerve fibers arises from the posterior end of the ganglion. The trunk extends to the fins and during its passage gives off nerves to the middle and posterior portions of the mantle.
Many of the features discussed in the foregoing sections exhibit peculiarities in their structure and position. However, until all the families of the Oegopsida are better understood anatomically, it is impossible to evaluate the systemic significance of these features.
Growth
Position of viscera (Pl. V, Fig. A; Pl. VI, Fig. A, D). One of the most remarkable features of Batoteuthis skolops is the position of the viscera in the far anterior region of the mantle cavity, with the exception of the gonad, which is near the posterior end of the mantle.
The relative location of different internal organs undergoes drastic change during ontogeny. However, the gills seem to maintain a constant position within the mantle cavity. In the youngest specimen available (Elt. 1314, 30-mm mantle length) the visceral organs are exceedingly small and are concentrated in a small area dorsal to the posterior base of the funnel and well anterior to the bases of the gills (Pl. VI, Fig. D) . The liver is small and oval. The stomach and ceacum lie in the midline anterior to the branchial hearts.
In holotype (Eltanin, 1396, 51-mm mantle length) the liver is greatly enlarged and the rest of the organs have been displaced posteriorly during growth (Pl. VI, Fig. A), bringing the stomach-caecum near the bases of the gills and posterior to the branchial hearts, which have remained in approximately the same position as in the smaller specimen.
The branchial hearts of the larger specimen (Elt. 382, 90-mm mantle length) have migrated well posterior to the bases of the gills (Pl. V, Fig. A). Although the stomach and caecum are missing, it is probable that they too would occur somewhat posterior to their former position.
Tentacular clubs. The club of the holotype occupies 80 % of the tentacle length. In the smaller specimen, however, the fully developed portion of the club occupies about 55 % of the tentacle length, but the sucker-bearing portion continues proximally along the stalk (36% of the tentacle length) , where the suckers occur in progressively fewer rows and become more scattered. Apparently this segment of the tentacular stalk serves as the zone of proliferation of club suckers.
Etymology
The generic name is derived from the Greek "batos" meaning thorn-bush. The one term combines two distinctive features of the genus: the long thornlike tail and the large bushy clubs. The Greek word "skolops" concerns anything pointed, again referring to the long pointed tail of the species.
Types
The type specimen is deposited in the United States National Museum. Holotype, USNM 576029.
Type Locality
South Pacific Ocean approximately 1000 km southeast of Wellington, New Zealand, at 49°57'S, 169º01'W in the region of the Antarctic Convergence. Eltanin Sta. 1396.
Distribution
This species has been captured in the South Atlantic and South Pacific oceans between approximately 49º and 60°S and between 55° and 170°W.
The vertical distribution is difficult to ascertain with certainty because of the use of open nets. The specimens were taken in 3-m Isaacs-Kidd Midwater Trawls, which fished at depths of 366, 1257, 1866, and 2525 meters.”
(Young & Roper, 1968: 185-197.