nomes no trilho de navegação
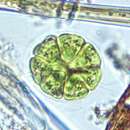
The Volvocaceae are a family of unicellular or colonial biflagellates, including the typical genus Volvox. The family was named by Ehrenberg in 1834,[1] and is known in older classifications as the Volvocidae. All species are colonial and inhabit freshwater environments.
The simplest of the Volvocaeans are ordered assemblies of cells, each similar to the related unicellular protist Chlamydomonas and embedded in a gelatinous matrix. In the genus Gonium, for example, each individual organism is a flat plate consisting of 4 to 16 separate cells, each with two flagella. Similarly, the genera Eudorina and Pandorina form hollow spheres, the former consisting of 16 cells, the latter of 32 to 64 cells. In these genera each cell can reproduce a new organism by mitosis.[2]
Other genera of Volvocaceans represent another principle of biological development as each organism develops differented cell types. In Pleodorina and Volvox, most cells are somatic and only a few are reproductive. In Pleodorina californica a colony normally has either 128 or 64 cells, of which those in the anterior region have only a somatic function, while those in the posterior region can reproduce; the ratio being 3:5. In Volvox only very few cells are able to reproduce new individuals, and in some species of Volvox the reproductive cells are derived from cells looking and behaving like somatic cells. In V. carteri, on the other hand, the division of labor is complete with reproductive cells being set aside during cell division, and they never assume somatic functions or develop functional flagella.[2]
Thus, the simplest Volvocaceans are colonial organisms but others are truly multicellular organisms. Larger volvocaceans have evolved a specialized form of heterogamy called oogamy, the production of small motile sperm by one mating type and relatively larger immotile eggs by another. Among the Volvocaceans are thus the simplest organisms with distinguishable male and female members. In all Volvocaceans, the fertilization reaction results in the production of a dormant diploid zygote (zygospore) capable of surviving in harsh environments. Once conditions have improved the zygospore germinates and undergoes meiosis to produce haploid offspring of both mating types.[2]
Colony inversion during development is a special characteristic of this order that results in new colonies having their flagella facing outwards. During this process reproductive cells first undergo successive cell divisions to form a concave-to-cup-shaped embryo or plakea composed of a single cell layer. Immediately after, the cell layer is inside out compared with the adult configuration—the apical ends of the embryo protoplasts from which flagella are formed, are oriented toward the interior of the plakea. Then the embryo undergoes inversion, during which the cell layer inverts to form a spheroidal daughter colony with the apical ends and flagella of daughter protoplasts positioned outside. This process enables appropriate locomotion of spheroidal colonies of the Volvocaceae. The mechanism of inversion has been investigated extensively at the cellular and molecular levels using a model species, Volvox carteri.[3]
Spheroidal colony inversion evolved twice during evolution of the Chlamydomonadales. In the Volvocaceae inversion first occurred when the Volvocaceae diverged from the closely related Goniaceae (see figure). It also occurred during evolution of Astrephomene. Inversion differs between the two lineages: rotation of daughter protoplasts during successive cell divisions in Astrephomene, and inversion after cell divisions in the Volvocaceae. [3]

The Volvocaceae are a family of unicellular or colonial biflagellates, including the typical genus Volvox. The family was named by Ehrenberg in 1834, and is known in older classifications as the Volvocidae. All species are colonial and inhabit freshwater environments.
Volvocaceae es una familia de algas perteneciente al orden Volvocales.
Volvocaceae es una familia de algas perteneciente al orden Volvocales.
Les Volvocaceae sont une famille d'algues vertes de l'ordre des Chlamydomonadales selon AlgaeBase (1er avril 2018)1er_avril_2018_1-0">1er_avril_2018-1">[1], selon ITIS (1er avril 2018)1er_avril_2018_2-0">1er_avril_2018-2">[2] et selon NCBI (1er avril 2018)1er_avril_2018_3-0">1er_avril_2018-3">[3] mais des Volvocales selon World Register of Marine Species (1er avril 2018)1er_avril_2018_4-0">1er_avril_2018-4">[4].
Le nom vient du genre type Volvox qui vient du latin volvo, « je roule » (infinitif volvere « rouler, tourner »), en référence à la forme en boule de l'algue[5]. La marque d'automobiles suédoises Volvo a la même étymologie.
Selon AlgaeBase (1er avril 2018)1er_avril_2018_1-1">1er_avril_2018-1">[1] :
Les Volvocaceae sont une famille d'algues vertes de l'ordre des Chlamydomonadales selon AlgaeBase (1er avril 2018), selon ITIS (1er avril 2018) et selon NCBI (1er avril 2018) mais des Volvocales selon World Register of Marine Species (1er avril 2018).
Volvocaceae, porodica zelenih alga u redu Chlamydomonadales ili Volvocales. Ime je dobila po rodu Volvox, a postoji 67 priznatih vrsta[1]
Volvocaceae, porodica zelenih alga u redu Chlamydomonadales ili Volvocales. Ime je dobila po rodu Volvox, a postoji 67 priznatih vrsta
Volvocaceae Ehrenberg, 1843 è una famiglia di alghe verdi appartenenti all'ordine delle Volvocales.[1]
Volvocaceae Ehrenberg, 1843 è una famiglia di alghe verdi appartenenti all'ordine delle Volvocales.
Maurakuliniai (lot. Volvocaceae) – žaliadumblių (Chlorophyta) šeima, priklausanti maurakuliečių (Volvocales) eilei.
Lietuvoje randamos šios gentys:
Volvocaceae é uma família de organismos biflagelados, unicelulares ou coloniais, que incluem o género típico, Volvox. O nome da família foi atribuido em 1834, por Ehrenberg in 1834. Em classificações mais antigas é conhecida como Volvocidae.[1]
Volvocaceae é uma família de organismos biflagelados, unicelulares ou coloniais, que incluem o género típico, Volvox. O nome da família foi atribuido em 1834, por Ehrenberg in 1834. Em classificações mais antigas é conhecida como Volvocidae.
Во́львоксовые (лат. Volvocaceae) — семейство зелёных водорослей. Представители обладают одноклеточным или ценобиальным строением. Клетки с двумя жгутиками.
Семейство насчитывает около 60 видов в 15 родах[1]:
Во́львоксовые (лат. Volvocaceae) — семейство зелёных водорослей. Представители обладают одноклеточным или ценобиальным строением. Клетки с двумя жгутиками.

本文参照
ボルボックス科 Volvocaceae は、群体性の緑藻類の群。決まった数の細胞が群体を作り、鞭毛を持って泳ぎ回る。群体がゼラチン質に包まれているのが特徴。
ボルボックス科の藻類は、典型的な細胞群体を作る。個々の細胞はクラミドモナスに似たもので、葉緑体を持ち、光合成を行う。また2本の等長鞭毛を先端に持っている。
群体の形は属によって決まっているが、多くは細胞が放射状に等距離に並び、それがゼラチン質に包まれたほぼ球形のものである。細胞は群体の外向きに鞭毛を出し、これによって回転しながら遊泳する。ボルボックスの「Volvo」はラテン語で「回転する」という意味である。例外はゴニウム属で、細胞は平面に並び、鞭毛は片方の面と外側に向かう。ただしこの属は現在はこの科から外されている。
群体を作る細胞は基本的には同じ構造を持つが、はっきりした分化を見せる例がある。栄養細胞として分化が見られ、群体に前後の区別がある例や、生殖細胞が特に分化する例などが知られ、ボルボックスは細胞数で最大であるほかに、この方向でももっとも特化したものである。
無性生殖時には群体を構成する細胞が分裂を繰り返し、決まった細胞数の小さな群体を作って独立する形で繁殖する。有性生殖では、同形配偶子接合、異形配偶子接合、卵生殖まである。異形配偶子を作る場合、小配偶子は1個の細胞の分裂によって多数が形成され、形成された精子が束になる精子束を形成するのが特徴である。
なお、科の和名としては岡田他(1965)はオオヒゲマワリ科、水野(1964)はヒゲマワリ科を採っている。ここでは混乱を避けて学名カナ読みを採っておく。
個々の細胞は、クラミドモナスに似たものである。2本の等長鞭毛が先端側にある。細胞壁に代わり、ゼラチン質が細胞を包み、これが群体を構成する基盤となる。葉緑体は1個で、大きくて杯状をしており、ピレノイドが1個またはそれ以上ある。収縮胞は普通は2個、鞭毛の基部にある。またやはり鞭毛の基部に、普通は眼点が1個ある[1]。
全て複数の細胞が細胞を包むゼラチン質によって繋がった構造をしている。ゼラチン質は個々の細胞の回りにあって、隣の細胞のそれと区別出来る例と、その間に区切りが見えないものがある。細胞の配置はゴニウムで平面に並ぶのを例外に、それ以外の属では放射状に並んで球形に近い形になり、鞭毛を外側に向ける。鞭毛を使って回転しながら泳ぐことが出来る。群体の構造は放射方向にほぼ同じだが、進行方向はある程度決まっており、それに応じて細胞に分化が見られる例もある。
無性生殖は個々の細胞が分裂し、それぞれに新しい群体を形成する形で行われる。例えばユードリナでは成熟した群体のそれぞれの細胞が同時に分裂を始め、それぞれの位置で分裂を繰り返し、16個ないし32個に達すると分裂を止め、それらが新たな群体となる。ただ、この娘群体は形成当初は鞭毛を内側に向けており、その後一端から反転し、鞭毛を外側に向けることで完成する。この様な反転は、同じような細胞の配列を持つこの科の他の属のものでも知られている。なお、この様な形成過程を採るため、この類の群体を構成する細胞数は2の累乗の値を採る。また、細胞数は群体の成長の間、変化しない。典型的な細胞群体である。
有性生殖は、同形配偶子接合から異形配偶子接合、それに卵生殖に至る様々な段階のものがある。
精子が形成される場合、その元になる細胞が分裂を繰り返して精子を形成するが、この時に程長い紡錘形の優性配偶子が束になって形成される。これは見た目が菊の花のようで、これを精子束(sperm packet)と言い、これを一つの群体と見なすことも出来る。雌性配偶子は元になる細胞が分裂せずに作られる[2]。
単細胞生物と多細胞生物の違いの一つに細胞に分化が見られるかどうか、という点がある。その観点では、この群は簡単ながら細胞に分化が見られる例があり、興味深いものである。
分化は、栄養細胞にも、生殖細胞にも見られる。ゴニウム属、パンドリナ属などではそのいずれもがほとんど見られない。それでもパンドリナでは進行方向の細胞が後方の細胞より眼点が大きいという程度の差はある。この傾向がもっとも顕著なのがプレオドリナで、この属のものは進行方向に近い一部の細胞が、後方の細胞より明確に小さい。似た傾向はユードリナの一部にも見られる。逆にアストレフォメネ属では、後方に少数の小さい細胞を持つ。ボルボックスでは表面にある鞭毛細胞と、内側にあるゴニジアと呼ばれる細胞の分化があり、これは後述の通り生殖に関わるものである。ただしゴニジアの配置は群体の後方に多い[3]。
生殖細胞では、ゴニウムやパンドリナでは全ての細胞が同じように生殖に与る。それに対してプレオドリナやアストレフォメネでは小さい細胞が生殖に参加しない。さらにボルボックスでは、群体を構成する大部分の細胞である表面の鞭毛細胞は生殖に関わらない。内部に位置するゴニジアと言う細胞のみが生殖に参加する。
概して細胞数が多くなるほどに分化の程度が高くなる傾向があり、細胞数ではほぼ以下のような順になる。
この中でパンドリナまでが同形配偶子接合、ユードリナ以上外径配偶子、ボルボックスだけが不動の卵を形成する[4]。
細胞の構造からクラミドモナスとの関係が論じられてきた。現在検討されたものについては、その全てが Chlamydomonas reinhardtii と類縁が深いとの結果が出ている[5]。
ちなみにボルボックスについてはその形態が動物の胞胚に似ることから、ヘッケルによって多細胞動物の進化との関連を論じられたことがあるが、現在では否定されている[6]。ただし、海綿類には胞胚の段階で最初に内側に繊毛が形成され、その後に反転するものがあることが知られている[7]。
この群に類するものとしては以下のような属が代表的である。細胞数の少ない方から挙げる。
これらは典型的なものでは判別が容易だが、困難な例もある。例えばパンドリナには細胞の集合が弱いものがあり、その場合にユードリナと混同される。プレオドリナは幼い群体では細胞の大きさに差がなく、これらの場合に判別は混乱する。従って有性生殖を見なければ判断出来ないなどの例も生じる[8]。
伝統的にはこれら全てをボルボックス科に含めた。現在ではシアワセモ(とBasichamys)をテトラバエナ科に、ゴニウムとアストレフォメネ属をゴニウム科とする扱いが行われている。ボルボックス科には以下の7属が含まれる[9]。
他にクラミドモナス型の細胞が集まった細胞群体を作るものにスポンディロモルム(クミヒゲマワリ)類がある。個々の細胞がゼラチン質を持たず、房状に寄り集まった群体を形成するものである[10]。これも細胞群体であり、同じくボルボックス目に含めるが、系統は異なると考えられている。
他に黄金色藻類にもSynura (シヌラ・モトヨセヒゲムシ)のように放射状に鞭毛細胞が集まった群体を作るものはある[11]。

