nomes no trilho de navegação
Els lumbrícids (Lumbricidae) són una família de cucs de terra que inclou la majoria d'espècies d'Europa. Unes 33 espècies de lumbrícids s'han naturalitzat per tot el món,[1] però la majoria són holàrtiques i d'Euràsia al Japó. N'hi ha unes 670 espècies dins 42 gèneres.[2]
Els lumbrícids (Lumbricidae) són una família de cucs de terra que inclou la majoria d'espècies d'Europa. Unes 33 espècies de lumbrícids s'han naturalitzat per tot el món, però la majoria són holàrtiques i d'Euràsia al Japó. N'hi ha unes 670 espècies dins 42 gèneres.
Die Regenwürmer (Lumbricidae) sind im Erdboden lebende, gegliederte Würmer aus der Ordnung der Wenigborster (Oligochaeta). Sie gehören innerhalb des Stammes der Ringelwürmer (Annelida) zur Klasse der Gürtelwürmer (Clitellata). Weltweit waren 2008 etwa 670 Arten der Regenwürmer (der Familie Lumbricidae) bekannt.[1] Zusätzlich wird aber eine unbekannte Anzahl morphologisch nicht unterscheidbarer Kryptospezies vermutet.[2]
In der Schweiz und in Deutschland leben derzeit 46 Arten,[3] in Österreich 62.[4] Nicht alle der in Europa lebenden Arten sind ursprünglich dort heimisch. Ihre durchschnittliche Lebenszeit liegt zwischen drei und acht Jahren. Der 9 bis 30 Zentimeter lange Tauwurm oder Gemeine Regenwurm (Lumbricus terrestris, früher auch als vermis terrae[5] bezeichnet) ist neben dem 6 bis 13 Zentimeter langen Kompostwurm (Eisenia fetida) wohl die bekannteste einheimische Annelidenart.
Während die Zuordnung in Europa relativ eindeutig ist, gibt es in anderen Erdteilen zahlreiche weitere Anneliden anderer Familien, die eine vergleichbare Lebensweise besitzen und auch Regenwürmer (oder earthworms) genannt werden. Die gesamte Gruppe ist untereinander verwandt und wohl monophyletisch. Sie wird nach einem morphologischen Merkmal, dem mehrlagigen Clitellum, Crassiclitellata genannt. Sie umfasst neben den Lumbriciden die Familien Acanthodrilidae, Megascolecidae, Octochaetidae, Ocnerodrilidae, Eudrilidae, Hormogastridae, Lutodrilidae, Ailoscolecidae, Sparganophilidae, Glossoscolecidae, Kynotidae, Almidae und Microchaetidae, wobei einige dieser Familien in ihrer Abgrenzung und Eigenständigkeit taxonomisch umstritten sind. Wird ein Tier als „Regenwurm“ bezeichnet, kann es also, außerhalb Europas, auch einer anderen Familie der Crassiclitellata angehören.
Die Herkunft der Bezeichnung „Regenwurm“ ist umstritten. Einer Ansicht zufolge soll er auf den althochdeutschen Begriff „Regnwurm“ zurückgehen, der sich auf das Verhalten der Würmer beziehe, bei starken Regenfällen die unterirdischen Wohnröhren rasch zu verlassen, um auf der Erdoberfläche dem Wasseranstieg im Oberboden zu entkommen. Nach anderer Ansicht rührt der deutsche Name von ihrer steten unterirdischen Aktivität her; noch im 16. Jahrhundert soll es die Bezeichnung „reger Wurm“ gegeben haben.
Auf den eigentlichen Aufenthaltsort des Wurms bezogen sind dagegen beispielsweise die englische Bezeichnung „earthworm“ („rainworm“ ist falsch und existiert nur als Dialektausdruck[6]) das türkische „yer solucanı“ und der französische Begriff „ver de terre“ (Erdwurm).
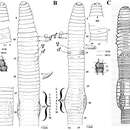
Der Körper des Regenwurms besteht aus zahlreichen zylindrischen Gliedern (Segmenten), die außen an ihren Seiten die kaum aus der Haut hervorragenden Borsten tragen. Die Borsten, von denen Regenwürmer pro Segment vier Paare besitzen, bestehen aus Chitin und Proteinen und können mit Hilfe besonderer Muskeln bewegt werden. Die Anzahl der Segmente nimmt mit dem Alter des Wurms zu. Eine spezielle Wachstumszone in der Nähe des Hinterendes produziert neue Glieder. Ausgewachsene Exemplare erreichen um die 160 Segmente.
Nach außen hin ist der gesamte Körper des Wurms und damit auch jedes seiner Segmente durch einen Hautmuskelschlauch abgegrenzt. Auf eine einschichtige Epidermis, die einige Drüsen- und Sinneszellen enthält und nach außen von einer kollagenhaltigen Cuticula umgeben ist, folgt eine Ringmuskelschicht. An diese schließt wiederum nach innen die dicke Längsmuskelschicht an. Die meisten Arten besitzen Hautpigmente. So sind zum Beispiel viele Lumbricus-Arten mehr oder weniger rot gefärbt. Alle Allolobophora-Arten besitzen dagegen mehr dunkle Pigmente, die die Hautoberfläche eher hellgrau oder grau-schwarz erscheinen lassen.
Eine Art Oberlippe, auch Kopflappen (Prostomium) genannt, überwölbt am Kopfende den Mund. Die Mundöffnung führt in den Darm, der den Regenwurm von vorn bis hinten vollständig durchzieht. Der Darm beginnt mit dem muskulösen Pharynx, auf den die Speiseröhre (Oesophagus) mit ihren Kalksäckchen sowie ein muskulöser Kropf und Muskelmagen folgen. Hier wird (ähnlich wie bei Hühnern) die pflanzliche Nahrung durch mitaufgenommene kleine Steinchen (hier Sandkörner) gleichmäßig zerrieben. Es folgt der lange Mitteldarm, der auf der Rückenseite in seiner gesamten Länge eine Einstülpung (Typhlosolis) aufweist, die die innere Darmoberfläche vergrößern hilft. Am Hinterende des Wurms befindet sich der After. Mit Hilfe kalziumhaltiger Abscheidungen neutralisieren die Würmer alle aufgenommenen säurehaltigen Bodeninhaltsstoffe und sorgen so auf natürliche Weise für eine Verbesserung des pH-Werts im Oberboden.
Zwischen den inneren Organen und dem Hautmuskelschlauch liegt in jedem Segment rechts und links ein mit Flüssigkeit ausgefüllter und von einer elastischen zarten Haut umgebener Hohlraum, die sekundäre Leibeshöhle (Coelom). Die abgrenzenden Quer- und Längswände in den Segmenten werden als Dissepimente bzw. Mesenterien bezeichnet. Die eingeschlossene Flüssigkeit wirkt für den wirbellosen Organismus als hydrostatisches Skelett. Im Zusammenwirken mit dem Hautmuskelschlauch unterstützt es den Wurm sowohl beim Bohren im Boden als auch bei der Fortbewegung allgemein, z. B. beim Kriechen an der Bodenoberfläche oder in der Wohnröhre.
Die Ausscheidungsorgane beginnen hinten in jedem Coelomsäckchen der Segmente (mit Ausnahme der ersten drei Glieder und des letzten Segments) links und rechts vom Darm mit je einem Wimperntrichter der sogenannten Nephridien. Diese gehen im nächsten Coelomsäckchen in einen langen, in Schleifen gewundenen, von Blutgefäßen umsponnenen Exkretionskanal über, der sich im Endabschnitt zu einer Harnblase erweitert. Der Wimpertrichter saugt Coelomflüssigkeit an, deren größter Teil im Exkretionskanal ins Blut transportiert wird (bis auf die Abfallstoffe). Die Adern dienen gleichzeitig der Versorgung des Ausscheidungsorgans mit Sauerstoff und Nährstoffen. Diese Versorgung der Zellen ist nötig, weil an den Schleifen des Nephridiums aktive Transportvorgänge zur Ausscheidung von Harnsäure, Harnstoff, Ammonium und Salzen sowie zur Resorption von Wasser, Ionen und organischen Verbindungen ablaufen.
Regenwürmer besitzen als Zwitter sowohl weibliche als auch männliche Geschlechtsorgane und diese zudem jeweils beidseitig. Sie beginnen mit zwei Paar Hoden jeweils vorn im Coelom der Segmente 10 und 11, angeheftet an deren vordere Scheidewände (Mesenterien oder Septen). Die Hoden sind bei einigen Gattungen in eine Samenkapsel eingeschlossen, bei anderen frei. Die reifen Spermien werden in einem Vorratsbehälter (Vesikel) gesammelt, der sich als Ausstülpung des hinteren Dissepiments in das folgende Segment erstreckt. Die Spermien werden von paarigen Spermienleitern (vasa deferentia), jeweils mit einem Wimperntrichter (einer Öffnung mit einem Saum von Zilien, die in Richtung des Inneren schlagen) aufgefangen, diese vereinigen sich beiderseits und münden meist in zwei erkennbaren Öffnungen (männliche Poren) nach außen. Die männlichen Genitalporen liegen meist im 13. oder 15. Segment. Im Segment 13 liegen entsprechend die weiblichen Organe (Eierstöcke). Die Eier werden von Eileitern, ebenfalls mit trichterförmiger Öffnung ins Coelom des Segments zu porenförmigen Öffnungen im folgenden 14. Segment geleitet. Bei der Paarung werden die Spermien des jeweiligen Partners zunächst in Samentaschen (Receptacula seminis) aufgenommen, deren Anzahl gattungsspezifisch verschieden ist. Die Befruchtung erfolgt im durch das Clitellum gebildeten Kokon (vgl. im Abschnitt Fortpflanzung).
Das Nervensystem ist hoch entwickelt. Es ist in das Gehirn oder Oberschlundganglion, das Bauchmark und die Segmentalnerven untergliedert. Das aus zwei miteinander verwachsenen Cerebralganglien bestehende Gehirn liegt im dritten Segment kurz vor dem Beginn des Pharynx dorsal dem Darm auf. Von ihm ziehen zahlreiche Nerven nach vorn in Richtung Prostomium. Schlundkonnektive verbinden das Oberschlundganglion auf beiden Seiten des Vorderdarms mit dem zu Beginn des vierten Segments ventral vom Darm gelegenen Unterschlundganglion (siehe nebenstehende Abbildung). Es folgt der Hauptstrang des Nervensystems, der auf der Bauchseite den Wurm vom vierten Kopfsegment bis zum Schwanzsegment durchzieht. Er wird daher als Bauchmark bezeichnet. In einem Frontalschnitt durch das Bauchmark erkennt man, dass es sich evolutionär vom Strickleiternervensystem ableitet. Die ursprüngliche Organisation des Strickleiternervensystems besteht aus paarigen, längs zur Körperachse verlaufenden Konnektiven, die durch quer zur Körperachse angeordnete Kommissuren miteinander verbunden sind. Konnektive und Kommissuren sind durch Nervenknoten (Ganglien), die überwiegend aus den Zellkörpern der Nervenzellen bestehen, miteinander verbunden. Beim Regenwurm sind diese Elemente alle in einem median verlaufenden (unpaaren) Nervenstrang vereint. In azangefärbten histologischen Präparaten kann man die beiden Faseranteile (Kommissuren und Konnektive) sowie die Nervenknoten auf geeigneten Schnitten gut differenzieren.
Pro Segment zweigen vom Bauchmark je drei Paar Segmentalnerven ab. Das vordere Paar liegt im kranialen (kopfwärts orientierten) Abschnitt eines Segments; das mittlere und das hintere Paar liegen meist eng benachbart im caudalen (schwanzwärts orientierten) Bereich eines Segments. Diese typische Anordnung erlaubt in den allermeisten Fällen, ein histologisches Präparat nach kopfwärts/schwanzwärts zu orientieren. Nach ihrer Abzweigung vom Bauchmark verlaufen die Segmentalnerven zunächst durch die sekundäre Leibeshöhle des Regenwurms (Coelom) und treten dann in den Hautmuskelschlauch ein, wo sie sich in einen ventralen und einen dorsalen Ast auftrennen. In ihrem weiteren Verlauf zwischen Ring- und Längsmuskulatur zweigen fortwährend feine Fasern ab und innervieren die Muskelzellen sowie die Zellen der Epidermis.
Die Innervation der Dissepimente, also der muskulösen Scheidewände zwischen den Segmenten, erfolgt durch die sogenannten Septalnerven, die in den Achseln der vorderen Segmentalnerven vom Bauchmark abzweigen.
Das Darmnervensystem des Regenwurms, das man auch stomodaeales System nennt, wurde erst relativ spät entdeckt. Augen fehlen zwar, doch ist der Regenwurm vor allem am Vorder- und Hinterende lichtempfindlich. In der Epidermis finden sich einzelne Sehzellen. Somit ist der Regenwurm zumindest in der Lage, hell und dunkel zu unterscheiden.[7] Der Regenwurm reagiert sehr sensibel auf Erschütterungen des Bodens.
Besondere Atmungsorgane besitzt der Regenwurm nicht, aber ein vielfach verzweigtes, geschlossenes Blutgefäßsystem, das den über die Haut aufgenommenen Sauerstoff und die aus dem Darm aufgenommenen Nährstoffe im ganzen Körper verteilt. Es besteht aus einem Rückengefäß, das das Blut von hinten nach vorn treibt, und einem Bauchgefäß. In den Segmenten sieben bis elf werden die beiden Hauptblutgefäße durch muskulöse und stark kontraktile Schlingen, die sogenannten Lateralherzen (zwei pro Segment), miteinander verbunden. In den übrigen Segmenten gibt es zwischen dem Rücken- und dem Bauchgefäß keine direkte Verbindung über Ringgefäße, sondern über Seitengefäße, die sich in den Organen und im Hautmuskelschlauch weiter zu Kapillaren verzweigen und dann wieder vereinigen.[8] Das Blut selbst ist durch den roten Blutfarbstoff Hämoglobin, der im Blutplasma gelöst ist, rot gefärbt. In den Hautkapillaren gibt das Blut Kohlendioxid an die Luft ab und nimmt Sauerstoff auf, der an das Hämoglobin gebunden wird, sich teilweise aber auch direkt im Wasser des Blutes löst (Hautatmung); umgekehrt verläuft dieser Gasaustausch in den Kapillaren der inneren Organe. Um die Hautatmung zu gewährleisten, muss die Haut feucht bleiben. So kann der Regenwurm auch in sauerstoffreichem Wasser überleben.[9] Das Blut enthält auch farblose Blutkörperchen, die Amoebocyten, die jedoch meistens den Gefäßwänden anliegen. Das Hämoglobin des Regenwurms besteht nicht wie das des Menschen aus nur 4, sondern aus 24 Untereinheiten. Entsprechend hoch ist die molare Masse von 3.840.000 g·mol−1.
Die Regenwürmer der Familie Lumbricidae haben wie die meisten Crassiclitellata (Regenwürmer im weiteren Sinne) zylindrische Körper, manchmal mit einem abgeflachten Körperende, bisweilen auch mit einem viereckigen, achteckigen oder trapezoiden Körperquerschnitt. Die meisten Lumbriciden besitzen Dorsalporen, kleine Öffnungen am Rücken, in denen das Coelom nach außen mündet und mit denen Wasser abgegeben oder aufgenommen werden kann.
Die Lumbriciden haben keinen Kaumagen im Bereich des Oesophagus, dafür aber einen Kaumagen im Mitteldarm, der 1 bis 2 Segmente zwischen dem 17. und dem 20. Segment einnimmt. Davor sitzt ein Kropf in einem einzigen Segment zwischen dem 15. und dem 17. Segment. Die Tiere haben in ihrer Vorderdarmwand Kalkdrüsen in einigen oder allen Segmenten zwischen dem 10. (manchmal auch 9.) und dem 15. Segment. Die großen Nephridien sind wohl entwickelt.
Das Clitellum der Lumbriciden ist meist sattelförmig und nimmt 4 bis 32 Segmente zwischen dem 17. und dem 52. Segment ein. Es weist zudem eine bandförmige oder papillenartige Tubercula pubertatis auf. Entsprechend dem Grundbauplan der Crassiclitellata haben die Lumbriciden stets zwei Paar Hoden im 10. und 11. Segment (holandrisch), dahinter aber nur ein Paar Eierstöcke im 13. Segment (metagyn), von denen die beiden Eileiter zu dem bauchseitig gelegenen Paar weiblicher Geschlechtsöffnungen am 14. Segment führen. Die Spermienleiter dagegen vereinigen sich, von den vier Hoden kommend, zu einem Paar, kreuzen mehrere Segmente, darunter das eierstocktragende, und führen so zum Paar der männlichen Geschlechtsöffnungen, das sich hinter den weiblichen bauchseitig am 15. Segment, selten am 13. oder 14. Segment noch vor dem Clitellum befindet. Die paarigen, adiverticulaten (keine Blindsäcke aufweisenden) Receptacula seminis sind entweder interparietal oder breiten sich frei ins Coelom aus und münden über paarige Öffnungen in 2 bis 8 Furchen zwischen den Übergängen vom 5. zum 6. Segment und vom 19. zum 20. Segment nach außen, meist 2 Paar Öffnungen in den beiden Furchen zwischen dem 9. und dem 11. Segment.
Regenwürmer verfügen über ein beachtliches Regenerationsvermögen. So ist es den Tieren möglich, nach der Durchtrennung ihr Hinterende fast vollständig wieder auszubilden, wobei das Regenerationsvermögen zur Körpermitte hin abnimmt. Am Vorderende können je nach Art maximal die ersten vier bis acht Segmente und das vor der Mundöffnung gelegene Prostomium abgetrennt werden. Diese Segmente werden wieder ersetzt. Die Regeneration des im 3. Segment gelegenen Gehirns (Oberschlund- und Unterschlundganglion) wurde beim Regenwurm Dendrobaena veneta (syn. Eisenia hortensis) untersucht. Fehlt das Gehirn, ist die Bildung der Gameten gehemmt, doch wird sie durch Regeneration des Gehirns wieder hergestellt. Die Gonaden (Hoden im 10. und 11. Segment, Eierstöcke im 13. Segment) können nach Verlust jedoch nicht wieder gebildet werden.[10][11] Trennt man vorn mehr Segmente ab, werden nicht mehr alle regeneriert. Bei mehr als 15 entfernten Segmenten ist meist keine Regeneration des Vorderendes mehr möglich. Das Regenerat hebt sich durch seine hellere Färbung von der benachbarten Körperpartie deutlich ab.
Das weit verbreitete Gerücht, dass zwei lebende Würmer entstünden, wenn man einen Wurm in der Mitte durchtrennt, trifft nicht zu. Jedes Körpersegment besitzt die genetische Anlage, den After wieder auszubilden, nicht aber den Kopf. Gelegentlich entsteht bei manchen Arten nach der Durchtrennung aus dem abgeschnittenen Hinterende ein Wurm mit zwei Aftern, der dann je nach Anzahl der noch verbliebenen Segmente über kurz oder lang verhungert.[12] Das Vorderende kann nur überleben, wenn die Teilung des Regenwurms hinter dem 40. Segment erfolgt. Davor befinden sich lebenswichtige Organe wie die Lateralherzen, die für die Aufrechterhaltung des Blutkreislaufs erforderlich sind und bei Verlust zum Tode führen. Auf Grund von Wundinfektionen überleben in freier Natur eher wenige durchtrennte Regenwürmer.[13]
Während der Regeneration fallen die Regenwürmer in eine Körperstarre. Dies machen sich Maulwürfe zunutze, die in die vordersten Segmente beißen um die dann unbeweglichen Regenwürmer in Kammern als Vorrat zu lagern.[14]
Die Würmer sind auch in der Lage, sich in bestimmten Gefahrensituationen selbst zu verstümmeln (Autotomie), z. B. wenn sie ein Fressfeind gepackt hat. Hierbei schnürt der Wurm am Hinterende eine Reihe von Segmenten ab und überlässt sie dem Räuber, um sich mit dem restlichen Körper durch Flucht in Sicherheit zu bringen.
Die Regenwürmer lassen sich in drei Gruppen einteilen:
Regenwürmer können anoxische Bedingungen (ganz ohne Sauerstoff) in wassergesättigten Böden nur bis zu etwa zwei Tage lang überstehen. Sie fehlen deshalb in grundwassergesättigten Böden. Auch nasse und dabei saure Moorböden werden nicht besiedelt. Typische Regenwurmarten wassergesättigter Böden sind Octolasium tyrtaeum und Proctodrilus antipae.[15]
Obwohl überwiegend terrestrisch, gibt es auch einige aquatische Regenwürmer, die stehende und fließende Gewässer besiedeln. Die einzige in Europa häufige und verbreitete Regenwurmart, die aquatisch und in wassergesättigten Böden lebt, ist Eiseniella tetraedra.[16] Diese Art kommt zum Beispiel verbreitet in der Gewässersohle des Rheins vor.[17]
Die nachtaktiven Regenwürmer sind überwiegend Substrat- und Pflanzenfresser. Das heißt, sie füllen ihren Darm mit humusreicher Erde und vermodertem Pflanzenmaterial. Sie ziehen nachts beispielsweise Keimlinge und Blätter in die Erde, um sie dort verrotten zu lassen und später als Nahrung zu verwerten. Um die Blätter festzuhalten, können Regenwürmer ihr Vorderende knopfartig aufblähen, sodass ihr Mund wie von einer Saugscheibe umgeben ist. Diese wird an das Blatt oder den Blattstiel gepresst und mit Hilfe des muskulösen Pharynx saugt sich der Wurm so sehr fest, dass er in der Lage ist, das angesaugte Blatt rückwärts kriechend in seine Wohnröhre zu ziehen. Sekrete aus den Pharynxdrüsen fördern den Zersetzungsprozess. Die aufgenommene Nahrung wird anschließend mit Hilfe des Muskelmagens zerrieben und im Mitteldarm verdaut.
Ständig fressen sich die Regenwürmer kreuz und quer durch die Bodenschichten ihres Lebensbereiches. Die dabei aufgenommene Erde enthält Detritus-Bestandteile, Bakterien, Pilzsporen und zahlreiche Einzeller, die verdaut und als Nahrung genutzt werden können. Manche Arten verzehren auch Aas. Durch die Beschaffenheit der Erde, die der Regenwurm erzeugt, wenn er die mitgefressenen Bodenbestandteile wieder ausgeschieden hat, werden die für den Boden nützlichen Mikroorganismen gefördert und die bodenfeindlichen eingedämmt, z. T. sogar vernichtet.
Mit Hilfe seiner Borsten und der Ring- und Längsmuskulatur ist der Regenwurm in der Lage, sich sowohl vorwärts als auch rückwärts kriechend zu bewegen. Sind beispielsweise beim Kriechen die Borsten schräg nach hinten gerichtet, bewirkt das Zusammenziehen der Ringmuskeln des Vorderendes, dass dieses dünner und länger wird. Dabei verankern die Borsten die hinteren Segmente im Boden, während der vordere Teil über den Boden gleitend sich nach vorne schiebt. Nun folgt eine von vorn nach hinten verlaufende Kontraktion der Längsmuskeln, wodurch die Segmente wieder dicker und kürzer werden, was den Wurmkörper nach vorne zieht (peristaltische Bewegung). Berührungs- und Lichtreize können Regenwürmer auch zu sehr raschen Muskelkontraktionen im Sinne einer Fluchtreaktion veranlassen.
Bei ihren Wanderungen durch die Böden bilden Regenwürmer Röhren. In lockerem Bodensubstrat wie zum Beispiel feuchten Waldböden oder Komposterde haben die Tiere beim Durchdringen des Bodens keine Probleme. Mineralböden dagegen bieten je nach Körnung, Festigkeit und aktuellem Wassergehalt sehr unterschiedliche Widerstände. Beim Eindringen in den Oberboden sowie beim Bau neuer unterirdischer Wohnröhren wird das verdünnte Vorderende als Bohrinstrument benutzt. Zum Überwinden des Bodenwiderstandes dient der stabile hydrostatische Druck der Leibeshöhlenflüssigkeit.
Meist werden die gebohrten Röhren mit Schleim und Exkrementen der Würmer ringsherum ausgekleidet und somit für den raschen Auf- und Abstieg stabilisiert. Man nennt diese Verfestigung auch „Tapete“. Sie dient auch den Pflanzen als Dünger.
Die Geschlechtsreife, die mit ein bis zwei Jahren eintritt, zeigt sich durch die Ausbildung des sogenannten Gürtels (Clitellum), einer gelblichen sattelförmigen, drüsenreichen Verdickung vom 27. bis 35. Segment. Die Seitenränder des Clitellums treten als sogenannte Pubertätsleisten besonders hervor.
Regenwürmer sind Zwitter und begatten sich wechselseitig. Große Drüsen des Gürtels scheiden bei der Begattung ein Sekret aus, mit dem sich jeder Wurm an der Bauchseite des 10. Segmentes des anderen Partners gegenläufig anheftet. Dann scheidet jeder Wurm aus den beiden Spermienleitern eine deutlich sichtbare Spermienportion aus, die er durch Hautbewegungen längs zweier Samenrinnen in Richtung seines Gürtels zu den dort befindlichen Samentaschen (Receptacula seminis) des Partners transportiert. Die dort gespeicherten fremden Spermien dienen ein paar Tage später zur Befruchtung der eigenen Eizellen. Bei einigen Regenwurmarten wurde hin und wieder auch Selbstbefruchtung beobachtet.
Die Eier werden (wie auch bei den Blutegeln) in Kokons abgelegt. Ein Clitellum-Sekret dient zur Bildung der Hülle dieses Ei-Kokons, ein zweites füllt es mit einer Eiweißschicht. Dann zieht sich der Wurm rückwärts aus dem Kokonring, in den dabei aus den Eileitermündungen je nach Art ein oder mehrere Eier und aus den Samentaschen Spermien abgegeben werden. Die Embryonen ernähren sich von dem Eiweiß, von dem sie umgeben sind, und machen im Ei nach einer kurzen Trochophora-Phase die Metamorphose zum zunächst durchsichtigen Wurm durch.
Die Entwicklungsdauer der Jungwürmer kann je nach Art und Umgebungstemperatur sehr verschieden sein. So schlüpft der Kompostwurm (Eisenia fetida) in seiner relativ warmen Umgebung bereits nach 16 bis 20 Tagen, dagegen benötigt Lumbricus terrestris bei einer mittleren Bodentemperatur von etwa 12 °C bis zu 135 Tage.
Das Temperaturoptimum der meisten Regenwurmarten liegt bei 10 bis 14 °C (Kompostwürmer: 20–25 °C). Darüber hinaus brauchen sie feuchte Erde. Dadurch sind Regenwürmer im Sommer und Winter weniger bis gar nicht aktiv.[18]
Die Wintermonate (Dezember bis Februar) verbringen Regenwürmer in Mitteleuropa in 40 bis 80 cm Bodentiefe in einer Art Kältestarre. Häufig finden sich unter wärmespeichernden Bodenstrukturen wie Baumstümpfen, Steinen oder Komposthaufen ganze Kolonien zusammengerollter Würmer. Unter hohen und dicht geschlossenen Schneedecken ist der Boden gegen Kälte geschützt und meist nicht gefroren. Stellenweise kann man hier Regenwürmer beobachten, die selbst im Winter im Bereich des Oberbodens aktiv sind. Noch ist unbekannt, inwieweit und wie lange die Tiere Kältegrade überstehen können. Mittelfristig droht den im Winter aktiven Würmern die Gefahr auszutrocknen, da eine Durchfeuchtung des Bodens aufgrund der gefrorenen Schneedecke bzw. Bodenoberfläche nicht stattfindet. Manche Arten können während der Winterruhe ca. 80 % ihres ursprünglichen Gewichts einbüßen, bevor sie sterben. Lumbricus terrestris zum Beispiel vollzieht in den relativ milden Gegenden Südwestdeutschlands (Oberrheingraben) keine richtige Winterruhe. Er erscheint in feuchten, frostfreien Nächten stets an der Bodenoberfläche, um Nahrung aufzunehmen.
Die im Herbst abgelegten Kokons der geschlechtsreifen Regenwürmer entwickeln sich im frostfreien Boden über den Winter hinweg weiter. Im Frühjahr schlüpfen die Jungwürmer nach Eintritt einer Bodentemperatur von über 10 °C.
Weshalb die Regenwürmer bei Regen ihre Wohnröhren verlassen, ist noch nicht vollständig geklärt. Oft wird angenommen, dass die Regenwürmer nicht an die Erdoberfläche kriechen, weil sie das feuchte Milieu der Niederschläge lieben, sondern weil sie bei Regen, insbesondere bei langanhaltenden Regenperioden, in ihren Gängen im Erdboden ersticken würden, da der im Wasser gelöste Sauerstoff nicht ausreicht, um den Wurm über die Hautatmung mit genügend Sauerstoff zu versehen. Wissenschaftliche Untersuchungen zeigten jedoch, dass selbst unter anaeroben Bedingungen unter Wasser gehaltene Regenwürmer erst nach 35 Stunden langsam zugrunde gehen. Wie sich weiter herausstellte, schalten die Würmer unter diesen extremen Bedingungen auf einen glykolytischen Stoffwechsel ohne Sauerstoffverbrauch um (Milchsäuregärung). Regenwürmer sind imstande, Überschwemmungen mit sauerstoffreichem Wasser bis zu mehrere Monate lang zu überleben. Tatsächlich erreichen sie in oft überschwemmten Auenbereichen sogar ungewöhnlich hohe Siedlungsdichten.[19]
Eine Studie der Carleton Universität im kanadischen Ottawa legt hingegen nahe, dass Regenwürmer aus Angst vor Maulwürfen an die Oberfläche kriechen: In Amerika ist es üblich, dass Angler Regenwürmer mittels „worm grunting“ (Substratschall, erzeugt durch einen in den Boden getriebenen Stock und eine vibrierende Metallscheibe) aus dem Boden austreiben, das Geräusch ähnle dem Grabegeräusch der Maulwürfe, wichtiger Fressfeinde von Regenwürmern.[20] Fallende Regentropfen sollen ähnliche Frequenzen erzeugen, was die Würmer eventuell mit grabenden Maulwürfen verwechseln. Eine zweite Studie in Florida erzielte vergleichbare Ergebnisse.[21] Ob diese Ergebnisse, erzielt an den amerikanischen Diplocardia mississippiensis, Diplocardia floridana und Pheretima diffringens, Familie Acanthodrilidae, allerdings allgemein gültig sind, ist unklar. Lumbricidae scheinen auf Substratschall nicht in vergleichbarer Weise zu reagieren.[22]
Regenwürmer können in bestimmten Bereichen einen Anteil von bis zu 90 Prozent der Biomasse der gesamten Bodenfauna ausmachen, wobei die Wurmdichte bis zu 2000 Individuen pro Quadratmeter erreichen kann.[23] Sie nehmen als Destruenten eine zentrale Stellung beim Abbau organischer Substanzen ein. Die lufthaltigen Gänge der Regenwürmer sorgen dafür, dass aerobe Bakterien mit genügend Sauerstoff versorgt werden und sich abgestorbene Pflanzenteile besser zersetzen.
Die gezielte Verarbeitung von Kompost (Kompostierung) durch Regenwürmer (Wurmkompost) ergibt als Produkt den sogenannten Wurmhumus mit hochkonzentrierten Bestandteilen an pflanzenverfügbaren Nährstoffen.
In den Llanos genannten Ebenen in Kolumbien und Venezuela entstanden vor allem durch Ausscheidungen von Regenwürmern der Gattung Andiorrhinus (Familie Glossoscolecidae) gebildete, Quadratkilometer große Landschaften aus bis zu fünf Meter breiten Erdhügeln, genannt Surales.[24]
Ihren Kot setzen die Regenwürmer meist oberirdisch in Form von geringelten Kotbällchen am Mündungsende ihrer Gänge ab. Schon 1881 beschrieb Charles Darwin in seinem Buch Die Bildung der Ackererde durch die Tätigkeit der Würmer die Tatsache, dass Regenwürmer beständig die aus den tieferen Schichten des Bodens stammende Erde durch ihren Darm hindurch an die Erdoberfläche befördern und dadurch zur Auflockerung und Belüftung der Böden beitragen.[25] Als begleitender Effekt zeigt sich das erleichterte Eindringen von Wasser in tiefere Bodenschichten. Dies wiederum fördert das Pflanzenwachstum. In den vertikal gebohrten Gängen können aber auch Pflanzenwurzeln schneller in die Tiefe wachsen.
Nach Darwins Berechnung befördern die Regenwürmer in vielen Teilen Englands jährlich auf einem sechs Hektar großen Landstück ein Gewicht von mehr als 25.000 kg Erde an die Oberfläche und bewirken dadurch eine ganz erhebliche Durchmischung der Bodenschichten, wobei der Untergrund mit Humusstoffen angereichert wird. Durch diese Tätigkeit „versinken“ Steine oder auch verlorene Münzen im Boden, weil die Ausscheidungen, die auf der Oberfläche abgelagert werden, aus tieferen Bodenschichten stammen. Dieser Vorgang wird allgemein als Bioturbation bezeichnet. In den Oberböden der Tropen und Subtropen wurden noch wesentlich höhere Umsetzungsraten festgestellt. Es liegt nahe, dass die Böden des tropischen Regenwaldes hierbei an der Spitze liegen (bis zu 280 t pro ha).
Ebenfalls 1881 hatte aber Louis Pasteur auch darauf hingewiesen, dass in den Kotbällchen der Regenwürmer Krankheitserreger aus tieferen Erdschichten an die Oberfläche gelangen können. Damals war es üblich, an Milzbrand-Erregern (Bacillus anthracis) verstorbene Rinder, Schafe und Pferde auf einer Acker- oder Wiesenfläche zu vergraben. In den Kotbällchen der Regenwürmer über diesen Kadavern waren hohe Konzentrationen an Milzbrand-Erregern entdeckt worden, die nachweisbar von Weidetieren aufgenommen wurden und zu weiteren Infektionen führten.[26] Ein Jahr später erörterte auch Robert Koch diese Form der Infektion in seiner Publikation „Über die Milzbrandimpfung“.[27]
Im Freiland sind die positiven Einflüsse von Regenwürmern nicht messbar, da man sie von den anderen Umwelteinflüssen nicht trennen kann. Unter standardisierten Bedingungen im Labor hingegen sind die Auswirkungen ihrer Tätigkeiten auf die Bodenverbesserung belegbar.
Für den biologischen Gartenbau sind Regenwürmer von zentraler Bedeutung. Regenwürmer gelten als wichtigste Erzeuger von Dauerhumus, gleichbedeutend mit den Ausscheidungen des Regenwurms, einer stabilen Bodenstruktur, ideal für das Pflanzenwachstum und mit vielen für die Pflanzen verfügbaren Nährstoffen. Daher ist auch die Pflege des Bodens in Form von Abdecken oder oberflächliches Hacken gegen Austrocknung, Mulchen und Einbringen von Kompost eine Vergünstigung der Lebensbedingungen für das Bodenleben (Edaphon) und somit für die Regenwürmer.
Der Komposthaufen im biologischen Gartenbau stellt sozusagen die Verdauungstätigkeit des Regenwurms im großen Stil nach. Hier finden sich vor allem der Kompostwurm und der Rote Waldregenwurm sehr häufig ein, ebenso wie unter ausgebrachtem Mulchmaterial. Die Reife des Kompostes lässt sich dadurch feststellen, dass der Haufen zusammengesunken ist und die Regenwürmer diesen verlassen haben.
Die Nährstoffanreicherung durch die Regenwürmer wird indirekt durch organische Düngung erzeugt und auf Kunstdünger wird explizit verzichtet. Da die Grabetätigkeit der Regenwürmer den Boden ausreichend lockert, ist im biologischen Garten bei richtiger Bodenpflege ein Umgraben im Gegensatz zur konventionellen Anbaumethoden nicht mehr erforderlich.
Regenwürmer dienen zahlreichen Vogelarten als Nahrungsquelle. Meist sind es Stare, Drosseln und Krähen, im Norden auch vermehrt Möwen und Austernfischer, die den Würmern gezielt nachstellen. Weitere natürliche Feinde sind Marder, Maulwürfe, Igel, Spitzmäuse, Erdkröten, Frösche, Feuersalamander, Hundertfüßer, Ameisen, Laufkäfer und Landplanarien. Auch Füchse und Dachse ernähren sich gern von Regenwürmern.
Rotkehlchen mit Regenwurm
Kaskadenfrosch (Amolops sinensis) beim Verzehr eines Regenwurms
Laufkäfer (Carabus auratus) beim Fang eines Regenwurmes
Schwarzer Moderkäfer attackiert einen Regenwurm
Maulwürfe beißen den Regenwürmern häufig ins Vorderende, um sie am Wegkriechen zu hindern. Die auf diese Art und Weise fluchtunfähig gewordenen, aber noch lebensfähigen Würmer werden anschließend an einem sicheren Platz unter der Erde als Nahrungsvorrat deponiert, zum Beispiel für die Wintermonate.
In Regenwürmern leben zahlreiche parasitierende Organismen. Neben verschiedenen, zum Teil symbiotisch lebenden Bakterien, Ciliaten und Flagellaten finden sich besonders häufig Gregarinen (Sporozoen) und Fadenwürmer (Nematoden). Befallen werden vor allem die Leibeshöhle sowie die Samenblase. Die meisten Parasiten sind harmloser Natur, einige aber übertragen als Zwischenwirte schwere Krankheiten (zum Beispiel die Lungenwurmkrankheit bei Schweinen und Hühnern durch Metastrongylus-Arten). Hin und wieder werden auch Larven von Bandwürmern (Eucestoda) in Regenwürmern nachgewiesen. Gelegentlich parasitieren auch Larven der Goldfliege (Lucilia sericata) in Regenwürmern. Sie halten sich bevorzugt im vorderen Bereich des Regenwurms (drittes und viertes Segment) auf und führen nach einiger Zeit zum Tod ihres Wirts.
Über die Gefährdung einzelner Regenwurmarten ist vergleichsweise wenig bekannt, weil in den meisten Regionen die Fauna schlecht bekannt ist. Selbst für das faunistisch gut erforschte Deutschland liegt erst seit 2013 ein Rote Liste vor, die im Rahmen einer Projektarbeit am Senckenberg Museum für Naturkunde Görlitz erstellt worden ist. Von den 47 hier etablierten Arten gelten 14 Arten als extrem selten, 2 Arten sind im Bestand gefährdet.[28] Zahlreiche gefährdete endemische Arten mit kleinem Verbreitungsgebiet sind von der Balkanhalbinsel bekannt.[29] Global betrachtet ist, neben der ackerbaulichen Nutzung von früheren Wäldern, die Einschleppung exotischer Arten mit Verdrängung der lokalen Fauna einer der wesentlichen Gefährdungsfaktoren. Besonders bedroht ist etwa die artenreiche Fauna der karibischen Inseln.[30]
Dichte und Häufigkeit von Regenwürmern hängen von Bodenfaktoren und Landbewirtschaftung ab. Regenwürmer sind selten oder fehlen ganz in sandigen Böden mit niedrigem pH-Wert. Außerdem hängt ihr Vorkommen von der Pflanzenstreu ab. Unter Nadelwald leben deshalb weniger Regenwürmer als unter Laubwald, besonders selten sind sie in Eucalyptus-Forsten.
Aufgrund des Einflusses auf die Bodenstruktur und die Streuzersetzung wird in landwirtschaftlichen Böden meist eine hohe Regenwurmdichte angestrebt. Extrem vermindert wird ihre Dichte allerdings durch Pflügen, worunter besonders die großen Arten spezifisch leiden. Während organische Düngung, z. B. Kompostgaben, günstig sind, vermindert Ausbringen von Gülle ihre Siedlungsdichte. Auch von vielen Pestiziden sind negative Auswirkungen nachgewiesen (insbesondere Fungizide und Insektizide). Für Herbizide mit dem Wirkstoff Glyphosat konnte eine im August 2015 erschienene Studie nachweisen[31], dass der Tauwurm (Lumbricus terrestris) nach Herbizidanwendung seine Aktivität fast völlig einstellte, während der Wiesenwurm (Aporrectodea caliginosa) unvermindert aktiv blieb. Die Herbizid-Anwendung reduzierte außerdem beim Wiesenwurm die Vermehrungsrate um 56 %[31]. Ungünstig sind insbesondere auch hohe Gaben kupferhaltiger Substanzen. Dies war lange Zeit aufgrund der geringen Empfindlichkeit des als Testorganismus eingesetzten, in natürlichen Böden nicht vorkommenden Kompostwurms Eisenia fetida unterschätzt worden.[32]
Die meisten Regenwurmarten können relativ einfach in Gefangenschaft gehalten und entsprechend gut vermehrt werden. Auf diese Weise werden Regenwürmer vielerorts in sogenannten Wurmfarmen in großem Stil gezüchtet und kommerziell genutzt. Vielfache Verwendung finden die Würmer als Futtertiere im Zoofachhandel oder als Köder für den Angler. Zuchtansätze und Zubehör zur Wurmzucht können von darauf spezialisierten Unternehmen im Internet bestellt und auf dem Postweg zugeschickt werden.
Seit einiger Zeit werden Wurmkulturen auch für die Bodenverbesserung und für die Kompostwirtschaft eingesetzt. Am besten eignen sich hierfür Arten, die bereits von Natur aus hohe Umsetzungs- und Reproduktionsraten aufweisen, z. B. Eisenia fetida.
Neuerdings werden auch tropische Regenwurmarten in geheizten Anlagen kultiviert, z. B. Eudrilus eugeniae aus Westafrika. Solche Arten sollten allerdings nur in geschlossenen Bereichen (Gewächshäusern, Laboreinheiten) gezüchtet werden. Ins Freiland ausgebracht werden sollten sie wegen der Neozoenproblematik jedoch nicht. Auch für den Hobbygärtner und den Halter von Terrarientieren (z. B. Schildkröten, Frosch- und Schwanzlurche) kann sich die Zucht von Regenwürmern in sogenannten Wurmkisten lohnen. Diese speziellen Behältnisse eignen sich u. a. auch für die Aufstellung auf Balkonen und Terrassen.
Immer wieder tauchen in der Presse Berichte auf, dass Regenwürmer für den menschlichen Verzehr gezüchtet und angeboten werden (z. B. als Fleischklößchen – sog. „Wormburger“ oder frisch frittiert). Aufgrund der generell starken Parasitierung der Würmer ist hier aber Vorsicht geboten (s. w. o. Parasiten).
Die Thielemannsche Oktettmethode[33] ist eine in der Wissenschaft inzwischen anerkannte Anwendung zum Fang von Regenwürmern mittels elektrischen Stroms. Das Verfahren nach dem Biologen Ullrich Thielemann wird häufig im Rahmen von Untersuchungen zur standardisierten Bestandserfassung der Regenwurmfauna spezieller Standorte angewandt. Auch im Zuge des Biomonitorings ist es eine weit verbreitete Nachweismethode. Hierbei stößt man acht Elektroden mit einem Abstand von etwa 50 cm zueinander kreisförmig in den Oberboden. Je nach Leitfähigkeit des anstehenden Bodens werden „zerhackte“ Gleichspannungsimpulse von 50 bis 250 Volt an die Elektroden für die Dauer von etwa 20 Minuten angelegt. Binnen weniger Minuten werden die im elektrischen Feld angesiedelten Regenwürmer aus dem Boden getrieben, wobei die größeren Exemplare meistens zuerst an die Oberfläche kriechen.
Eine andere und auch üblichere Methode für die Gewinnung von Regenwürmern aus dem Erdreich ist die Anwendung von Formaldehyd. Zur Gewinnung der Tiere werden 50 ml 37-prozentiges Formalin auf 10 Liter Leitungswasser vermengt und die so erzeugte Formalinmischung auf etwa 1/2 m² Rasen- oder Bodenfläche verteilt. Die so gewonnenen Tiere werden nach dem Auflesen zur Abschleimung ca. 10 bis 15 Minuten in sauberes Leitungswasser gelegt. Für die Bestimmung der Tiere z. B. unter einem Binokular werden sie nach einer Erholungspause von etwa zwei Stunden mit CO2-Gas betäubt. Danach werden die Würmer auf ein vorbereitetes Zuchtsubstrat gelegt, wo sie sich nach fünf bis zehn Minuten eingraben.
Aufgrund der potentiellen Auswirkungen des Formalins auf andere Bodenorganismen ist diese Methode nicht zu empfehlen.
Als Alternative zum giftigen und umweltschädlichen Formaldehyd wird seit Anfang der 1990er Jahre auch Senf als (zudem kostengünstigeres) Austreibungsmittel empfohlen. Bei dieser Methode werden zunächst 60 g Senfmehl in einen halben Liter Wasser gegeben. Nach einer Stunde Wartezeit und gründlicher Durchmischung wird die Suspension in 9,5 Liter Wasser gegeben. Ebenso kann Fertigsenf verwendet werden, der leichter zu handhaben ist, da er direkt in das Wasser eingerührt werden kann; jedoch ist hier die Austreibungswirkung im Hinblick auf bestimmte Regenwurmarten (namentlich der Gattungen Aporrectodea und Allolobophora) geringer.[34]
In bestimmten Regionen Kanadas, der USA und Englands werden Regenwürmer in großem Maße mit Vibrationen gejagt. Die Methoden nennen sich worm grunting, worm charming und worm fiddling.[35][36]
Die Familie Lumbricidae gehört in eine Klade der Gürtelwürmer (Clitellata), die nach einem morphologischen Merkmal, dem mehrlagigen Clitellum, als Crassiclitellata bezeichnet wird. Die Crassiclitellata entsprechen den Regenwürmern (oder engl. earthworms) im weiteren Sinne. Sie sind (fast) weltweit verbreitet, während Lumbricidae selbst nur holarktisch in Europa, Teilen Westasiens und Nordamerika vorkommen. Nach phylogenomischen Daten sind sowohl die Crassiclitellata wie auch die Lumbricidae monophyletisch.[37] Die Analyse der Verwandtschaftsverhältnisse war jahrzehntelang durch die morphologische Einförmigkeit der Tiere, ihr hohes evolutives Alter, verbunden mit hoher Evolutionsgeschwindigkeit in einigen Teilgruppen, behindert worden, wodurch zahlreiche frühere systematische und taxonomische Hypothesen später widerlegt wurden. So erwiesen sich etliche Gattungen als polyphyletische, nach einem Merkmal zusammengefügte Verlegenheits-Taxa, sind also keine evolutiven Einheiten.[38][39] Dies betrifft, mindestens, die „Gattungen“ Allolobophora, Aporrectodea, Cataladrilus, Cernosvitovia, Dendrobaena, Helodrilus und Octodrilus, möglicherweise auch weitere.
Innerhalb der Crassiclitellata wird eine Überfamilie Lumbricoidea unterschieden, die, in revidierter Fassung[37], die Familien Lumbricidae, Hormogastridae (unter Einschluss der monotypischen Ailoscolecidae)[40], Criodrilidae und Lutodrilidae umfasst. Am engsten verwandt, und vermutlich Schwestergruppen, sind die Familien Criodrilidae und Hormogastridae. Die Criodrilidae sind eine kleine Familie mit nur einer oder zwei Gattungen und vermutlich nur drei Arten, mit paläarktischer Verbreitung, von der europäischen Mittelmeerregion und Nordafrika bis Japan.[41] Die Hormogastridae umfassen vier Gattungen mit knapp 30 Arten, die an besonders trockene Böden adaptiert sind, und leben im westlichen Mittelmeerraum.[40]
Den genetischen Daten zufolge[39] entstand die Familie Lumbricidae schon in der Kreide. Die am frühesten abgespaltenen Arten leben im westlichen Mittelmeerraum und auf den Inseln Sardinien und Korsika, so dass eine Entstehung der Familie in Europa wahrscheinlich erscheint, wo sie auch heute ihren Verbreitungsschwerpunkt besitzt. Die nordamerikanischen Arten hätten den Kontinent noch vor der Spaltung des Urkontinents Laurasia besiedelt. Die ältesten Arten besitzen alle eine endogäische Lebensweise innerhalb des Erdbodens. Die epigäischen, in der Streuschicht lebenden und die anözischen, zwischen den Strata wechselnden Arten wären demnach jünger und von diesen abgeleitet.
Regenwürmer sind mit anderen als „Würmer“ bezeichneten Tieren nicht näher verwandt. Der Begriff der „Würmer“ kennzeichnet heute nur noch einen bestimmten Habitus, aber keine systematische Einheit.
In Deutschland wurden insgesamt 47[42] Regenwurmarten aus 15 Gattungen nachgewiesen, wobei mit dem Badischen Riesenregenwurm (Lumbricus badensis) nur eine endemische Art besteht. Die Anzahl der Arten nimmt von Norden nach Süden zu, 14 Arten kommen ausschließlich im Süden vor.[43]
In der Familie der Regenwürmer werden aktuell über 670 Arten in über 40 Gattungen anerkannt. Sie werden in ökologische Gruppen eingeteilt und umfassen nach der taxonomischen Einteilung der zuvor zitierten – auf Deutschland beschränkten – Untersuchung folgende Gattungen und Arten:
Gruppe der endogäischen Regenwürmer (wohnen in der oberen Mineralschicht des Bodens in horizontalen Gängen):
Gattung Allolobophora
Gattung Aporrectodea (syn. Allolobophora)
Gattung Helodrilus
Gattung Murchieona
Gattung Octodrilus
Gattung Octolasion
Gattung Proctodrilus
Gruppe der anecischen Regenwürmer (wohnen in oberen bis drei Meter tiefen Bodenschichten in vertikalen Gängen):
Gattung Aporrectodea
Gattung Fitzingeria
Gattung Lumbricus
Gruppe der epigäischen Regenwürmer (wohnen in organischer Auflage über dem Boden):
Gattung Allolobophoridella
Gattung Aporrectodea
Gattung Dendrobaena
Gattung Dendrodrilus
Gattung Eisenia
Gattung Eiseniella
Gattung Lumbricus
Gattung Satchellius
Zur Familie Lumbricidae gehören folgende Gattungen:
Regenwürmer finden auch in der Bioindikation Anwendung. Sie akkumulieren im Boden Schwermetalle derart, dass deren Aufnahme der Schwermetalle mit den entsprechenden Bodengehalten korreliert.[44] Das Monitoring von Regenwürmern dient vor allem zum Nachweis schleichender Bodenveränderungen.[45] Da Regenwürmer am Beginn einer Nahrungskette stehen, kommt ihnen aus ökologischer Sicht eine große Bedeutung zu.[44]
Für das Biomonitoring mit Regenwürmern wird aus einer Fläche von ungefähr einem halben Quadratmeter eine statistisch ausreichend große Anzahl von Regenwürmern eingesammelt. Die Probenahme ist jährlich zum gleichen Zeitpunkt zu wiederholen.[46]
Im Nordosten der Vereinigten Staaten wirkt sich das Ernährungsverhalten (vermutlich als Angelköder) eingeschleppter europäischer Regenwurmarten stellenweise negativ auf das Bodenökosystem der Laubwälder aus. Da dort seit der letzten Eiszeit keine Regenwürmer verbreitet waren, haben sich die Laubwälder darauf eingestellt, dass sich dichte Laubschichten auf dem Boden bilden, die im Winter als Isolierung dienen und kleinere Pflanzen sowie Baumschösslinge vor Frost schützen. Wo aber Regenwürmer – zum Beispiel der Tauwurm und der Rote Waldregenwurm – diese Schicht zersetzen, sind Böden und Unterwuchs dem strengen Frost des nordamerikanischen Winters ausgesetzt. Dies kann dort heimische Arten sowie die Waldverjüngung bedrohen.[47][48][49]
Ein Regenwurm ist der unkonventionelle Videospiel-Protagonist Earthworm Jim, ein schießfreudiger Superheld, der auch seine eigene Zeichentrick-Reihe erhielt. In der Zeichentrickserie „Die Biene Maja“ tritt ein Regenwurm mit Namen Max auf. Andere Videospiele, in denen Regenwürmer die Hauptrollen spielen, sind die der Worms-Reihe.
Die Regenwürmer (Lumbricidae) sind im Erdboden lebende, gegliederte Würmer aus der Ordnung der Wenigborster (Oligochaeta). Sie gehören innerhalb des Stammes der Ringelwürmer (Annelida) zur Klasse der Gürtelwürmer (Clitellata). Weltweit waren 2008 etwa 670 Arten der Regenwürmer (der Familie Lumbricidae) bekannt. Zusätzlich wird aber eine unbekannte Anzahl morphologisch nicht unterscheidbarer Kryptospezies vermutet.
In der Schweiz und in Deutschland leben derzeit 46 Arten, in Österreich 62. Nicht alle der in Europa lebenden Arten sind ursprünglich dort heimisch. Ihre durchschnittliche Lebenszeit liegt zwischen drei und acht Jahren. Der 9 bis 30 Zentimeter lange Tauwurm oder Gemeine Regenwurm (Lumbricus terrestris, früher auch als vermis terrae bezeichnet) ist neben dem 6 bis 13 Zentimeter langen Kompostwurm (Eisenia fetida) wohl die bekannteste einheimische Annelidenart.
Während die Zuordnung in Europa relativ eindeutig ist, gibt es in anderen Erdteilen zahlreiche weitere Anneliden anderer Familien, die eine vergleichbare Lebensweise besitzen und auch Regenwürmer (oder earthworms) genannt werden. Die gesamte Gruppe ist untereinander verwandt und wohl monophyletisch. Sie wird nach einem morphologischen Merkmal, dem mehrlagigen Clitellum, Crassiclitellata genannt. Sie umfasst neben den Lumbriciden die Familien Acanthodrilidae, Megascolecidae, Octochaetidae, Ocnerodrilidae, Eudrilidae, Hormogastridae, Lutodrilidae, Ailoscolecidae, Sparganophilidae, Glossoscolecidae, Kynotidae, Almidae und Microchaetidae, wobei einige dieser Familien in ihrer Abgrenzung und Eigenständigkeit taxonomisch umstritten sind. Wird ein Tier als „Regenwurm“ bezeichnet, kann es also, außerhalb Europas, auch einer anderen Familie der Crassiclitellata angehören.
Chincha llawq'a (Lumbricidae) nisqakunaqa chincha allpapachakunapi (chincha kuskanlunq'u nisqapi) allpa ukhupi kawsaq llawq'akunam.
Chincha llawq'a (Lumbricidae) nisqakunaqa chincha allpapachakunapi (chincha kuskanlunq'u nisqapi) allpa ukhupi kawsaq llawq'akunam.
De Mettjen (Lumbricidae), ok Maddiks, Madiks, Mecken, Meddiks oder Metten, in annere plattdüütsche Kuntreien ok Eulken, Pieren, Pierwörme, Purrewörmer un Uulewüpken nömmt, towielen ok, as hoochdüütsch Regenwörmer, sünd Wörmer ut de Ornen vun de Minnerbössen (Oligochaeta). Mank den Stamm vun de Ringelwörmer (Anelida) höört se to de Klass vun de Gördelwörmer (Clitellata). In dat Johr 2008 sünd up de ganze Eer bi 670 Aarden vun Mettjen (ut de Familie Lumbricidae) bekannt ween.[1] Tosätzlich warrt annahmen, datt dat noch en unbekannte Tahl vun Kryptospezies gifft, de sik in't Utsehn nich verscheelt, man nich to desülvigen Aarden hören doot.[2] In de Swiez un in Düütschland sünd hüdigendags 46 Aarden bekannt, in Belgien un de Nedderlannen man bloß 22,[3] in Öösterriek 54.[4] Nich all Aarden, de sik in Europa finnt, sünd dor al jummers tohuse ween. In'n Döörsnitt leevt se twuschen dree un fiev Johre lang. An'n bekanntesten sünd bi us de Normole Mettje (Lumbricus terrestris), de 9 bit 30 cm lang weern kann, un de Mestmettje (Eisenia fetida), de 6 -13 cm lang warrt.
In Europa is meist kloor to sehn, wenn en Worm to de Mettjen (Lumbricidae) tohören deit, man in annere Eerddeele gifft dat en grote Tahl vun Ringelwörmer ut annere Familien, de up lieke Aart leven doot un ok Mettjen (oder earthworms) nömmt weert. De ganze Grupp is unner’nanner verwandt un wohrschienlich monophyleetsch. Se warrt na en gemeensomen Kennteken in de Morphologie, dat is dat mehrlagige Clitellum, Crassiclitellata nömmt. Dor höört, blangen de Mettjen (Lumbriciden) noch de Familien vun de Acanthodrilidae, Megascolecidae, Octochaetidae, Ocnerodrilidae, Eudrilidae, Hormogastridae, Lutodrilidae, Ailoscolecidae, Sparganophilidae, Glossoscolecidae, Kynotidae, Almidae un Microchaetidae to. Bi en Reeg vun düsse Familien hefft sik de Forschers noch in' Plünnen, wie de in de Taxonomie intodelen sünd. En Deert, dat buten Europa „Mettje“ nömmt warrt, kann vundeswegen ok to en annere Familie mank de Crassiclitellata tohören.
De Mettjen (Lumbricidae), ok Maddiks, Madiks, Mecken, Meddiks oder Metten, in annere plattdüütsche Kuntreien ok Eulken, Pieren, Pierwörme, Purrewörmer un Uulewüpken nömmt, towielen ok, as hoochdüütsch Regenwörmer, sünd Wörmer ut de Ornen vun de Minnerbössen (Oligochaeta). Mank den Stamm vun de Ringelwörmer (Anelida) höört se to de Klass vun de Gördelwörmer (Clitellata). In dat Johr 2008 sünd up de ganze Eer bi 670 Aarden vun Mettjen (ut de Familie Lumbricidae) bekannt ween. Tosätzlich warrt annahmen, datt dat noch en unbekannte Tahl vun Kryptospezies gifft, de sik in't Utsehn nich verscheelt, man nich to desülvigen Aarden hören doot. In de Swiez un in Düütschland sünd hüdigendags 46 Aarden bekannt, in Belgien un de Nedderlannen man bloß 22, in Öösterriek 54. Nich all Aarden, de sik in Europa finnt, sünd dor al jummers tohuse ween. In'n Döörsnitt leevt se twuschen dree un fiev Johre lang. An'n bekanntesten sünd bi us de Normole Mettje (Lumbricus terrestris), de 9 bit 30 cm lang weern kann, un de Mestmettje (Eisenia fetida), de 6 -13 cm lang warrt.
In Europa is meist kloor to sehn, wenn en Worm to de Mettjen (Lumbricidae) tohören deit, man in annere Eerddeele gifft dat en grote Tahl vun Ringelwörmer ut annere Familien, de up lieke Aart leven doot un ok Mettjen (oder earthworms) nömmt weert. De ganze Grupp is unner’nanner verwandt un wohrschienlich monophyleetsch. Se warrt na en gemeensomen Kennteken in de Morphologie, dat is dat mehrlagige Clitellum, Crassiclitellata nömmt. Dor höört, blangen de Mettjen (Lumbriciden) noch de Familien vun de Acanthodrilidae, Megascolecidae, Octochaetidae, Ocnerodrilidae, Eudrilidae, Hormogastridae, Lutodrilidae, Ailoscolecidae, Sparganophilidae, Glossoscolecidae, Kynotidae, Almidae un Microchaetidae to. Bi en Reeg vun düsse Familien hefft sik de Forschers noch in' Plünnen, wie de in de Taxonomie intodelen sünd. En Deert, dat buten Europa „Mettje“ nömmt warrt, kann vundeswegen ok to en annere Familie mank de Crassiclitellata tohören.
Pierelinge (Lumbricidae) zeen 'n femielje van óngewerveldje bieëster die behuuere toete ringwörm (Annelida). Pierelinge behuuere toete Oligochaeta; dit is 'n groep van wörm die de zieë höbbe verlaoten en in zeutwater, mer ouch op 't landj kónne laeve. Pierelinge kómme weltwied gans algemein veur, oetgezunjerdj in druueg klimaten en op Antarctica.
Meistes zeen ze in groeate aantale te vinje; inkel saorte zeen weltwied oetgezatte gewaore veure baomstructuur te baetere. Pierlinge höbbe door 't grave van tunnels en 't aafbraeke van plantjaerdig matterjaal 'ne groeaten invlood oppe baomstructuur. Weltwied guuef 't óngevieër 670 saorten aan pierelinge die in lingdje variëre van inkel centimaeters toet decimaeters.
Bie 't trille vanne baom kroepe de pierelingen oete gróndj; dit kan dore loupbewaeginge van bieëster waere veroearzaak of door miensjelik hanjele, wie mit behölp van 'ne stek. Ónger netuurlike ómstenjigheje kómme pierelinge baove door graafbewaeginge van möl. Pierelinge kómmen ouch baovegróndjsj bie raengerechtig waer veur te paere; ze zeen wiejer veural 's nachs aktief. Inne wintjer kroepe ze deper de gróndj in en gaon ze in 'n saort van wintjerslaop of diapause. Sómmige gewerveldje bieëster zeen sterk aafhenkelik vanne pierelingen ómdet ze 'n wichtige brón van voor vörme; veurbiljer hievan zeen de mol enne maerelink.
Pierelinge (Lumbricidae) zeen 'n femielje van óngewerveldje bieëster die behuuere toete ringwörm (Annelida). Pierelinge behuuere toete Oligochaeta; dit is 'n groep van wörm die de zieë höbbe verlaoten en in zeutwater, mer ouch op 't landj kónne laeve. Pierelinge kómme weltwied gans algemein veur, oetgezunjerdj in druueg klimaten en op Antarctica.
Meistes zeen ze in groeate aantale te vinje; inkel saorte zeen weltwied oetgezatte gewaore veure baomstructuur te baetere. Pierlinge höbbe door 't grave van tunnels en 't aafbraeke van plantjaerdig matterjaal 'ne groeaten invlood oppe baomstructuur. Weltwied guuef 't óngevieër 670 saorten aan pierelinge die in lingdje variëre van inkel centimaeters toet decimaeters.
Bie 't trille vanne baom kroepe de pierelingen oete gróndj; dit kan dore loupbewaeginge van bieëster waere veroearzaak of door miensjelik hanjele, wie mit behölp van 'ne stek. Ónger netuurlike ómstenjigheje kómme pierelinge baove door graafbewaeginge van möl. Pierelinge kómmen ouch baovegróndjsj bie raengerechtig waer veur te paere; ze zeen wiejer veural 's nachs aktief. Inne wintjer kroepe ze deper de gróndj in en gaon ze in 'n saort van wintjerslaop of diapause. Sómmige gewerveldje bieëster zeen sterk aafhenkelik vanne pierelingen ómdet ze 'n wichtige brón van voor vörme; veurbiljer hievan zeen de mol enne maerelink.
Piern (Lumbricidae) beheurt tot de orde Haplotaxida, eleedte oligochaeetn.
Dr beent völ soortn piern. In Neerlaand en belgië al 25 soortn en oawer de hele wearld mear as 2200. Piern beent oaweral, behalve in dreuge gebeedn en Antarktika. De kleainste soortn wordt neet grötter as 2 cm, mear de grötste soortn köant wal 300 cm laank wordn en 8 mm dikke (Megascolides australis).
Sommige soortn goat recht noar euntern en heel deepe, zoas n reagnpier (Lumbricus terrestris), den at 9 tot 30 cm laank wordn kan, en de rooe pier (Lumbricus rubellis), den at 15 cm laank wordn kan. n Dauwpier blif kort oonder t oppervlak en löt heupkes driete achter an t eardoppervlak.
Dr beent ook piern dee at leawt in de streuiselloage, zo as n mestpier (Eisenia fetida). Disn wörd 6 tot 13 cm laank, en wördt um zinne rooe kluur en oranje strepe in Amerika ook wal tiegerwörm eneumd. Wieters keump n griesblaauwn Allolobophora caliginosa vake vuur.
In Humusoarme groond köant wal 62 piern eveundn wordn en in Humusrieke groond zitt mangs wal 432 piern per veerkaante meter.
As de groond trilt kroept de piern umhoge. Dit kan duur leu of deers veroorzaakt wordn, mear ook duur t wear, zo as ne gooe reagnbuje. Piern goat meestieds 's nachs gangs. Op n weenterdag kroept ze deeper de groond in en goat in n soort van weentersloap.
Nen pier besteet oet verskeaidene ringe (segmeantn). Oet iederen reenk stekt veer poar börstels, dee at met verskilnde spiern bewöagn köant wordn. Met at n pier eulder wördt, krig hee ook mear ringe, dee at evörmd wordt in n ampart stukn an t eande van t lief. An de vuurkaante, noa ongevear 30 ringe, zit bie volwassen piern ne verdikkige, ook wal t zadel (clitellum) eneumd.
An de boetnkaante van n pier zit ne hoedspierloage. t Is ne ééncellige loage van epidermis-celn, woerin at n poar tastzin-celn en kliern zitt. n Epidermis is of edekt met ne loage cuticula. Doaroonder zit ne loage dwars estreept spiervleais en doaroonder ne dikke loage glad spiervleais.
Piern hebt ne boawnlippe (prostomium) den an t kopeande oawer n bek haank. t Moondgat begeent bie n eerstn reenk en löp duur tot an n doarm, den at duur t hele lief van n pier löp. t Begin van n doarm hef nen spierlozen haals (farynx) den at oawer geet in n sloekdoarm (oesophagus). Disn sloekdoarm besteet oet kalkzekskes, dan n spierloozn krop en n stel stoarke maagn. Hierin wörd t plaantachtige voor met zaand liek vermoaln (zoas det ook bie henn geet, alleene doot dee t met kèjkes). Doarnoa keump n langn middeldoarm, den at oawer de hele achterkaante iets nen oetspoarige hef (typhlosolis), woerduur at hee van de binnkaante ne gröttere oppervlakte hef. An t eande zit t geatje van t gat.
Piern (Lumbricidae) beheurt tot de orde Haplotaxida, eleedte oligochaeetn.
Dr beent völ soortn piern. In Neerlaand en belgië al 25 soortn en oawer de hele wearld mear as 2200. Piern beent oaweral, behalve in dreuge gebeedn en Antarktika. De kleainste soortn wordt neet grötter as 2 cm, mear de grötste soortn köant wal 300 cm laank wordn en 8 mm dikke (Megascolides australis).
Sommige soortn goat recht noar euntern en heel deepe, zoas n reagnpier (Lumbricus terrestris), den at 9 tot 30 cm laank wordn kan, en de rooe pier (Lumbricus rubellis), den at 15 cm laank wordn kan. n Dauwpier blif kort oonder t oppervlak en löt heupkes driete achter an t eardoppervlak.
Dr beent ook piern dee at leawt in de streuiselloage, zo as n mestpier (Eisenia fetida). Disn wörd 6 tot 13 cm laank, en wördt um zinne rooe kluur en oranje strepe in Amerika ook wal tiegerwörm eneumd. Wieters keump n griesblaauwn Allolobophora caliginosa vake vuur.
In Humusoarme groond köant wal 62 piern eveundn wordn en in Humusrieke groond zitt mangs wal 432 piern per veerkaante meter.
As de groond trilt kroept de piern umhoge. Dit kan duur leu of deers veroorzaakt wordn, mear ook duur t wear, zo as ne gooe reagnbuje. Piern goat meestieds 's nachs gangs. Op n weenterdag kroept ze deeper de groond in en goat in n soort van weentersloap.
Reyðmaðkurin (frøðiheiti: Lumbricidae) blandar moldina og ger, at væta og luft seta niður í moldina og gera hana betri. Til eru fleiri enn 3000 reyðmaðkasløg. Hvussu fruktagóð moldin er, veldst nógv um reyðmaðkin. Sum maðkurin etur sær holur ígjøgnum moldina, koma ósodnaðar leivdir úr bakendanum. Flestir reyðmaðkar liva av ræ, men summir eru ránakendir og taka nógv ymisk smádýr. Hóast reyðmaðkur er liðormur, eru ikki allir liðirnir eins. Gøgn gongur gjøgnum alt dýrið. Nervalagið er í tveimum streingjum, ið liggja tætt saman nær undirborðinum og koma saman í fremra enda í einfaldan heila. Tá ið reyðmaðkur skríður, arbeiða æðrarnar sum hjarta.
Slėiks (luotīnėškā: Lumbricidae, onglėškā: Earthworms, vuokīškā: Regenwürmer) īr žėidouts kėrmėns, katros gīven pu žemė.
Slėikā gīven dirvuo, lėgė 10 cm gėloma. Žėimuos aba sausruos čieso gal īsėroustė 60–80 cm, sīkēs lėgė 2 m gėlomon. Jied vėsuokius pūviesius, trūniesius. Šliaužadoms pu žemė ana akie, poikėn dirvas viedėnėma, ondens nutekiejėma.
Lietovuos dirvūs vėinamė m³ žemės būn ~20–400 slėikū. Tonkiausē anėi gīven pėivūs, ganīklūs, ōžoulīnūs.
Slėikus jied kormē, šernas, paukštē. Žmuonis anūs tonkē kabėn kāp jouka žovaunont.
The Lumbricidae are a family of earthworms. About 33 lumbricid species have become naturalized around the world,[1] but the bulk of the species are in the Holarctic region: from Canada (e.g. Bimastos lawrenceae on Vancouver Island) and the United States (e.g. Eisenoides carolinensis, Eisenoides lonnbergi and most Bimastos spp.) and throughout Eurasia to Japan (e.g. Eisenia japonica, E. koreana and Helodrilus hachiojii). An enigmatic species in Tasmania is Eophila eti. Currently, 670 valid species and subspecies in about 42 genera are recognized.[2] This family includes the majority of earthworm species well known to Europeans.
The family consists of the following genera:
The worms in the family Lumbricidae originate from Europe, but over time members of the family have since been introduced and spread around the globe.[4]
Members of Lumbricidae are native to Europe and are most diverse in southern Europe.[4] There are 30 species from the family in Ireland and Britain. Notably, a single mature individual of the species Prosellodrilus amplisetosus was found in a survey of soil biodiversity in Ireland. P. amplisetosis had never been recorded in Ireland before and is commonly found in France or Spain. It is thought to have been introduced by humans through agricultural supplies.[5] Another interesting case is of the species Dendrobaena attemsi in Scandinavia. They were first found in a national park in Sweden, the furthest north the species have been found. The discovery of D. attemsi implies the range of the species is increasing north.[6] It is not only in Sweden that species of Lumbricidae are expanding their range. Many of the species found in Finland are exhibiting similar increases in range[7] and Lumbricidae worms are also expanding into Northeastern Europe, starting from the near Baltic sea.[8]
A 2022 molecular phylogenetic study of the highly diverse Franco-Iberian genus Zophoscolex showed most of the Iberian species to form a distinct clade, formally described as Castellodrilus stat. nov..[3] Other species were moved to the genera Cataladrilus and Compostelandrilus, with the remaining species remaining in Zophoscolex restricted to French representatives.[3]
Lumbricidae make up the majority of earthworms found in China, despite not being native to the area.[4]
At higher elevations in India, some species of Lumbicidae can be found.[4]
When European settlers came to North America, so did European earthworms like the Lumbricidae. Before this, the area in North America where glaciers had been were mostly worm-free.[9] Lumbricidae worms are known to be expanding into the Great Lakes region.[8] The introduced worms have an impact on the native species and environments. Species from the family, such as Lumbricus rubelles, are believed to have displaced the local species in a number of regions.[10] In others, Lumbricidae species outnumber the native species in terms of biomass. Despite this, they are not as productive, in terms of processing nitrogen and phosphorus, as the native species.[11] Lumbricidae worms also tend to have a higher species richness than native North American worms, though the species richness of both the native and Lumbricidae decreases with increasing latitudes.[12]
Similar to North America, worms from the family were introduced to New Zealand and Australia by European settlers.[13][4]
Harvestmen, especially from the genera Leiobunum and Hadrobunus, are known to consume Lumbricidae earthworms. This happens mostly in temperate regions.[14] Another species known to prey on Lumbricidae is the Bannan caecilian. Lumbricidae are an important part of its diet.[15]
The Lumbricidae are a family of earthworms. About 33 lumbricid species have become naturalized around the world, but the bulk of the species are in the Holarctic region: from Canada (e.g. Bimastos lawrenceae on Vancouver Island) and the United States (e.g. Eisenoides carolinensis, Eisenoides lonnbergi and most Bimastos spp.) and throughout Eurasia to Japan (e.g. Eisenia japonica, E. koreana and Helodrilus hachiojii). An enigmatic species in Tasmania is Eophila eti. Currently, 670 valid species and subspecies in about 42 genera are recognized. This family includes the majority of earthworm species well known to Europeans.
Los lumbrícidos (Lumbricidae), son una familia de lombrices de tierra, son anélidos oligoquetos del orden Crassiclitellata. Cabe destacar que el nombre de lombriz se aplica a otros invertebrados vermiformes sin relación alguna con los anélidos, como las lombrices intestinales (tenias, pertenecientes a los platelmintos) y Ascaris lumbricoides (del filo de los nematodos).
De origen acuático, mantienen aún características propias del medio: respiración cutánea, dependencia de la humedad, sistemas de resistencia a desecamiento (cápsulas, puestas masivas, quiescencia).[1]
Las lombrices más conocidas excavan galerías en el suelo y salen de noche a explorar sus alrededores. Son animales muy beneficiosos: mientras excavan para hacer sus túneles, ingieren partículas de suelo y digieren cualquier resto orgánico. En épocas húmedas arrastran hojas al interior de la tierra para alimentarse. Con ello remueven, airean y enriquecen el suelo, contribuyendo a que se mantenga fértil al hacer ascender fósforo y potasio del subsuelo y al expulsar sus propios desechos nitrogenados. La época más propicia para las lombrices es cuando el clima es húmedo y cálido, momento en el que salen a la superficie para procrear. Las lombrices son hermafroditas ya que poseen órganos reproductores masculinos y femeninos, pero necesitan aparearse (ver Oligochaeta).
Son muy voraces, llegando a comer hasta el 90% de su propio peso por día. De esta ingesta, excreta entre el 50 y 60%, convertido en un nutriente natural de altísima calidad, conocido como lombricompuesto o humus de lombriz.
Las lombrices de tierra no tienen dientes; su efectividad reside en su aparato digestivo para lo cual tiene un aparato bucal succionador, faringe, buche, molleja y el resto es intestino.
Su apareamiento se produce generalmente cuando asoman a comer a la superficie. La puesta de huevos se realiza a razón de un cocón por animal cada 45 a 60 días. Viven de 4 a 5 años. No posee pulmones: respira por la piel (respiración cutánea).
No pueden reproducirse asexualmente pero sí poseen capacidad de regeneración. Seccionadas en dos partes, el trozo de lombriz que contiene la cabeza regenera el extremo caudal.
Las lombrices de tierra tienen un importante interés práctico por muchos aspectos. Por ejemplo:
No cuenta con una cabeza bien definida pero sí con una boca en uno de sus extremos y un ano en el otro. No obstante puede moverse en ambos sentidos sin mayor problema.
El sistema muscular de la lombriz de tierra consiste en una serie de fibras externas circulares o transversas de músculo, que rodean el cuerpo, y una serie interna de fibras musculares longitudinales que sirven para mover las cerdas.
El aparato circulatorio está formado por un vaso sanguíneo dorsal prominente y por lo menos cuatro vasos sanguíneos ventrales, que recorren de forma longitudinal el cuerpo y están conectados entre sí a intervalos regulares por medio de una serie de vasos transversales. El vaso dorsal está equipado con válvulas y es el verdadero corazón. No obstante, el bombeo de la sangre se produce sobre todo por movimientos musculares generales.
El sistema nervioso central consiste en un par de ganglios suprafaríngeos, generalmente llamados cerebro, y un cordón ventral que pasa debajo del canal alimentario con ganglios en cada segmento. Las lombrices de tierra carecen de órganos sensoriales aparte de los del tacto. Pero sí podemos observar ya, en este organismo primitivo, los tres elementos esenciales del sistema nervioso: receptores, conductores y efectores; y es en el desarrollo de la cadena nerviosa en la lombriz de tierra donde los conductores quedan organizados en vías ascendentes y descendentes. En este caso, el sistema nervioso cuenta con tres tipos de células nerviosas especializadas.
El aparato digestivo está formado por una faringe musculosa, un delgado esófago, un buche o receptáculo de comida de paredes delgadas, una molleja muscular empleada para moler la tierra ingerida, y un intestino largo y recto.
Tienen un importante papel en los ecosistemas terrestres húmedos, donde son consideradas especies clave:
Allolobophora verde
Los lumbrícidos (Lumbricidae), son una familia de lombrices de tierra, son anélidos oligoquetos del orden Crassiclitellata. Cabe destacar que el nombre de lombriz se aplica a otros invertebrados vermiformes sin relación alguna con los anélidos, como las lombrices intestinales (tenias, pertenecientes a los platelmintos) y Ascaris lumbricoides (del filo de los nematodos).
De origen acuático, mantienen aún características propias del medio: respiración cutánea, dependencia de la humedad, sistemas de resistencia a desecamiento (cápsulas, puestas masivas, quiescencia).
Las lombrices más conocidas excavan galerías en el suelo y salen de noche a explorar sus alrededores. Son animales muy beneficiosos: mientras excavan para hacer sus túneles, ingieren partículas de suelo y digieren cualquier resto orgánico. En épocas húmedas arrastran hojas al interior de la tierra para alimentarse. Con ello remueven, airean y enriquecen el suelo, contribuyendo a que se mantenga fértil al hacer ascender fósforo y potasio del subsuelo y al expulsar sus propios desechos nitrogenados. La época más propicia para las lombrices es cuando el clima es húmedo y cálido, momento en el que salen a la superficie para procrear. Las lombrices son hermafroditas ya que poseen órganos reproductores masculinos y femeninos, pero necesitan aparearse (ver Oligochaeta).
Son muy voraces, llegando a comer hasta el 90% de su propio peso por día. De esta ingesta, excreta entre el 50 y 60%, convertido en un nutriente natural de altísima calidad, conocido como lombricompuesto o humus de lombriz.
Las lombrices de tierra no tienen dientes; su efectividad reside en su aparato digestivo para lo cual tiene un aparato bucal succionador, faringe, buche, molleja y el resto es intestino.
Su apareamiento se produce generalmente cuando asoman a comer a la superficie. La puesta de huevos se realiza a razón de un cocón por animal cada 45 a 60 días. Viven de 4 a 5 años. No posee pulmones: respira por la piel (respiración cutánea).
No pueden reproducirse asexualmente pero sí poseen capacidad de regeneración. Seccionadas en dos partes, el trozo de lombriz que contiene la cabeza regenera el extremo caudal.
Lierot (Lumbricidae) on harvasukasmatoihin kuuluva heimo, johon kuuluvia lajeja ovat muun muassa kasteliero ja tunkioliero.
Lierot (Lumbricidae) on harvasukasmatoihin kuuluva heimo, johon kuuluvia lajeja ovat muun muassa kasteliero ja tunkioliero.
Lumbricidae, les Lombricidés sont une famille de vers de terre caractéristique des régions tempérées de l'hémisphère nord et initialement originaire de la zone paléarctique (Europe, Afrique du Nord, Moyen-Orient et Asie).
Elle compose l'essentiel de la faune de vers de terre européenne. Introduite en Amérique du Nord avec l'arrivée des premiers Européens, elle s'est répandue jusqu'à composer aujourd'hui la quasi-totalité des espèces canadiennes à l'Est des Rocheuses. Elle a également été introduite dans les régions tempérées de l'hémisphère sud où elle prospère aujourd'hui (Australie, Afrique du Sud, Patagonie). Cette famille comprend plusieurs centaines d'espèces.
Ces espèces jouent un rôle majeur en termes de services écosystémiques, notamment par le travail du sol que les vers assurent, et par le rôle qu'ils jouent dans le cycle de la matière organique et du carbone dans le sol, ainsi qu'en matière de cinétique de métaux lourds et métalloïdes dans l'environnement (dans les sols pollués notamment)
Les Lombricidés ont été classé en 3 catégories écologiques par l'agronome Marcel Bouché dans les années 1970 en s’appuyant sur des critères morphologiques, physiologiques et écologiques[1] : les épigés, les endogés et les anéciques.
Les vers de terre épigés sont de petite taille et souvent de couleur brune pour se camoufler et se protéger des rayons lumineux. Ils vivent à la surface, dans des milieux riches en matière organique, notamment dans la litière dont ils se nourrissent et ne creusent pas ou très peu de galerie. Leur capacité de reproduction est très élevée, leur croissance rapide, leur mobilité importante et leur longévité courte[1],[2]. Ce sont notamment le Lombric châtain, Satchellius mammalis, le ver du fumier et Dendrobaena octaedra[3].
Les vers de terre endogés sont de taille petite à moyenne et de couleur claire ou dépourvus de pigmentation. Ils vivent continuellement dans les premiers niveaux du sol où ils produisent un réseau de galeries horizontales très ramifié et temporaire. Ils se nourrissent soit de racines mortes (saprophagie), soit de matière organique plus ou moins intégrée à la matière minérale (géophagie). Leur capacité de reproduction est moins rapide que celle des épigés et leur longévité plus importante[1],[2]. Ce sont par exemple la forme typique d'Aporrectodea caliginosa, Le Lombric rose, de couleur rosâtre ; le Lombric bleu et le Lombric marbré de couleur blanchâtre et la forme typique du Lombric vert, de couleur verdâtre[3].
Les vers de terre anéciques sont de taille moyenne à géante, et sont colorés de rouge à brun grisâtre. Ils vivent sur l'ensemble du sol où ils produisent un réseau de galerie verticales peu ramifié et permanent qui débouchent à la surface pour permettre à l'eau et à l'air de communiquer. Ils se nourrissent de la matière organique végétale de la surface et y déposent leurs déjections, nommées turricules. Leur capacité de reproduction est lente et leur longévité longue. Lors des périodes trop sèches et trop froides, ces espèces survivent grâce à une mise en inactivité nommée diapause ou quiescence[1],[2]. Ces espèces sont divisées en deux groupes selon leur morphologie. : les vers à tête rouge, au premier quart coloré de rouge à rouge pale, au clitellum orange et globalement trapu (ce sont par exemple le Lombric commun et Lumbricus rubellus rubellus) et les vers à tête noire, au premier quart coloré de noir à gris clair, au clitellum marron à marron clair et au pore male rosâtre bien visible à l’œil nu (par exemple Aporrectodea longa)[3].
Tous les Lombricidés sont originellement des hermaphrodites. Certains pratiquent une reproduction sexuée dite « bi-parentale » comme le Lombric commun. Plus précisément, ils sont hermaphrodites protandre, c'est-à-dire que chaque individu est alternativement mâle puis femelle. Deux vers mâles s'accouplent au niveau de leur clitellum (la bague colorée gonflée située au tiers antérieur du corps qui est la caractéristique générale des Anéciques), en contact tête-bêche. Après l'échange réciproque de leurs spermatozoïdes, ils les conservent et se transforment en femelle. Leur clitellum produit alors une capsule nommée cocon auquel ils adjoignent un ou plusieurs ovules qu'ils fécondent avec les spermatozoïdes de leur ancien partenaire. Ils font enfin glisser le cocon le long de la partie antérieure de leur corps jusqu'à ce qu'il s'en détache et se ferme à ses deux extrémités. Il est alors émis dans le sol. D'autres espèces pratiquent une reproduction asexuée, la parthénogénèse, à partir de leurs seules gamètes[4].
Les vers adultes produisent des quantité différentes de cocons par an, suivant leur âge, les conditions climatiques comme la sécheresse et le froid et leurs conditions de vie, notamment la pollution[5],[6]; des critères qui déterminent également la durée d’incubation[7],[8]. Des espèces anéciques ou endogées comme Aporrectodea caliginosa, le Lombric à tête noire et le Lombric bleu produisent entre 3 et 13 cocons par an alors que les épigés comme Lumbricus rubellus, le Lombric châtain et Dendrobaena rubidus sont capables d’en produire entre 42 et 106 par an[9]. Quant au Lombric commun, il peut produire entre 10 et 25 cocons par an en fonction des conditions de température[10]. Les cocons sont résistants à certaines conditions extrêmes comme la sécheresse, même si elle provoque leur déshydratation et retarde le développement de l'embryon. C'est notamment le cas pour Lumbricus rubellus, chez qui il n'est pas rare que seul le cocon survive aux estivations[11].
De ces cocons éclosent un ou plusieurs juvéniles, leur nombre par cocons étant variable en fonction de l'espèce : 20 % des naissances du Lombric bleu sont des jumeaux alors qu'elles ne sont que 1 % chez le Lombric commun et le Lombric à tête noire[12]. Chez le Lombric du fumier, un maximum 11 vers par cocon a été trouvé[13].
Le ver de terre juvénile acquiert d'abord des caractères sexuels sub-adultes comme le puberculum tuberculeux et les pores sexuels. Puis vient le caractère sexuel adulte typique, un organe lié au processus de ponte : le clitellum qui ressemble à une bague colorée gonflée située au tiers antérieur du corps. Mais ce temps de maturation est particulièrement variable d'une espèce à une autre et fonction des conditions de milieux comme la température, l'humidité et la nourriture disponible. Par exemple, un juvénile Aporrectodea caliginosa devient mature en 3 à 6 semaines[14]. Au champ, le Lombric commun devient mature généralement en 1 an[15] alors qu’il ne lui suffira que de quelques mois pour atteindre la maturité sexuelle en conditions de laboratoire[16].
La durée de vie des Lombricidés dépend de l’espèce, de leur biotope et des contraintes climatiques. Ainsi, le Lombric commun peut vivre plusieurs années en conditions de laboratoire[15] alors qu’en conditions naturelles et particulièrement dans les champs cultivés, son espérance de vie diminue à quelques mois[9]. Suivant le groupe, les stratégies varient entre les types « r » et les types « K », c'est-à-dire entre les espèces à durée de vie courte qui privilégient une reproduction prolifique et une croissance rapide, à savoir les épigés, et les espèces qui privilégient une durée de vie longue au détriment de la reproduction et de la croissance, à savoir les anéciques ; les endogés ayant une stratégie intermédiaire[17].
Les vers de terre et plus particulièrement les Lumbricidae sont des espèces ingénieures du sol car ils structurent le sol, stimulent l'activité microbienne, décomposent et brassent la matière organique tout en l'agrégeant aux éléments minéraux, aèrent et permettent une meilleure infiltration et rétention de eau. Ce réaménagement du sol, nommé bioturbation, a de nombreux bénéfices notamment la réduction de l’érosion, l’augmentation de la production végétale ainsi que la réduction des risques de pollution[18].
Du fait de leur localisation dans les couches supérieures du sol et de leur régime alimentaire, les vers de terre se trouvent parmi les premiers être vivants à être impactés par les polluants auxquels ils sont très sensibles. Ce sont par conséquent de très bon Bioindicateurs en rendant compte de l’état et des usages de l’écosystème que constitue le sol. C'est notamment le cas des populations vivant à proximité des zones industrielles et des zones urbaines qui sont contaminées par des métaux lourds comme le zinc, le cuivre, le cadmium, le mercure et le plomb[19],[20]. Les microplastiques représentent aussi une sérieuse menace pour les populations de vers de terre, surtout pour leur reproduction[6]. Au niveau agricole, les milieux voués à l'agriculture industrielle et pollués aux insecticides tels que les néonicotinoïdes représentent une menace directe pour les vers de terre et leurs prédateurs en s'accumulant également dans la chaîne alimentaire. À l'inverse, l'abondance des vers de terre augmente lorsque l’usage des pesticides diminue, les parcelles gérées en agriculture biologique étant plus favorables que celles en conventionnel[21],[22],[23].
Selon Ecobiosoil de l'Université de Rennes[2], et Termium plus[24] :
Selon World Register of Marine Species (13 janvier 2021)[25] :
Lumbricidae, les Lombricidés sont une famille de vers de terre caractéristique des régions tempérées de l'hémisphère nord et initialement originaire de la zone paléarctique (Europe, Afrique du Nord, Moyen-Orient et Asie).
Elle compose l'essentiel de la faune de vers de terre européenne. Introduite en Amérique du Nord avec l'arrivée des premiers Européens, elle s'est répandue jusqu'à composer aujourd'hui la quasi-totalité des espèces canadiennes à l'Est des Rocheuses. Elle a également été introduite dans les régions tempérées de l'hémisphère sud où elle prospère aujourd'hui (Australie, Afrique du Sud, Patagonie). Cette famille comprend plusieurs centaines d'espèces.
Ces espèces jouent un rôle majeur en termes de services écosystémiques, notamment par le travail du sol que les vers assurent, et par le rôle qu'ils jouent dans le cycle de la matière organique et du carbone dans le sol, ainsi qu'en matière de cinétique de métaux lourds et métalloïdes dans l'environnement (dans les sols pollués notamment)
Regenwormen (Lumbricidae) zijn een familie van ongewervelde dieren die behoren tot de ringwormen (Annelida). Regenwormen behoren tot de Oligochaeta, dit is een groep van wormen die de zee verlaten hebben en in zoetwater maar ook op het land kunnen leven. Regenwormen komen wereldwijd algemeen voor, uitgezonderd in droge klimaten en op Antarctica.
Meestal zijn ze in grote aantallen te vinden, enkele soorten zijn wereldwijd uitgezet om de bodemstructuur te verbeteren. Regenwormen hebben door het graven van tunnels en het afbreken van plantaardig materiaal een grote invloed op de bodemstructuur. Wereldwijd zijn er ongeveer 670 soorten regenwormen bekend,[1] die in lengte variëren van enkele centimeters tot decimeters.
Bij het trillen van de bodem kruipen de regenwormen uit de grond, dit kan door de loopbewegingen van dieren worden veroorzaakt of door menselijk handelen zoals met behulp van een stok of greep. Onder natuurlijke omstandigheden komen regenwormen boven door graafbewegingen van mollen. Regenwormen komen ook bovengronds bij regenachtig weer om te paren, ze zijn verder vooral 's nachts actief. In de winter kruipen ze dieper de grond in en gaan in een soort winterslaap of diapause. Sommige gewervelde dieren zijn sterk afhankelijk van de regenworm omdat ze een belangrijke voedselbron vormen, voorbeelden zijn de mol en de merel.
Regenwormen hebben een holarctische oorsprong: van Noord-Amerika tot oostwaarts Europa, Azië en het Midden-Oosten.[1] Sommige soorten hebben een wereldwijde verspreiding, zoals de gewone regenworm, en andere komen slechts in een bepaald gebied voor. De soort Eisenia japonica bijvoorbeeld is alleen te vinden in Japan. In Nederland en België komen 22 soorten voor, de bekendste soort is de gewone regenworm of pier (Lumbricus terrestris). Een lijst van alle Nederlandse soorten is rechts weergegeven in de uitklapbare lijst. De meeste van deze soorten hebben nog geen Nederlandse naam.
Regenwormen leven niet allemaal ondergronds, veel soorten zijn diepgravers die lange verticale gangen maken zoals de veel voorkomende dauwpier of gewone regenworm (Lumbricus terrestris), die 9 tot 30 cm lang wordt en de rode worm Lumbricus rubellus die tot 15 cm lang wordt. De gewone regenworm leeft vlak onder het grondoppervlak en laat hoopjes uitwerpselen achter op het bodemoppervlak. Er zijn er ook die in de strooisellaag leven zoals de mestpier Eisenia fetida, die 6 tot 13 cm lang wordt en door zijn rode kleur en soms oranje dwarsbanden in Amerika wel tijgerworm wordt genoemd. Verder komt de grijsblauwe Allolobophora caliginosa veel voor.
Hoewel de soorten verschillende voorkeuren hebben is toch een van de belangrijkste factoren de zuurgraad van de grond. De meeste regenwormen hebben een voorkeur voor een pH-neutrale tot iets zure (pH 5,4) grond. De soort Dendrobaena octaedra is echter nog aanwezig bij een pH van 4,3 en sommige vertegenwoordigers van de familie Megascolecidae hebben zich zelfs gespecialiseerd in extreem zure grond. Hoe zuurder de grond des te eerder gaan regenwormen in diapauze. Bij een pH van 6,4 blijven de regenwormen het langst in diapauze. Regenwormen kunnen niet overleven in een te zuur milieu, zoals veengronden. Hierdoor worden plantenresten niet op grote schaal omgezet in meststoffen, waardoor turf kan ontstaan.
Regenwormen hebben een langwerpig, typisch wormachtig lichaam zonder poten en ook andere aanhangsels ontbreken. De gewone regenworm bereikt een totale lichaamslengte van dertig centimeter, dit is aanmerkelijk langer dan gelijkende regenwormen zoals Lumbricus rubellus (tot 15 cm) en Lumbricus castaneus (tot 5 cm).[3] Sommige vertegenwoordigers van andere in de bodem levende wormen worden nog langer dan regenwormen, zoals Megascolides australis uit Australië die een lengte kan bereiken van drie meter. Deze soort wordt wel 'reuzenregenworm' genoemd maar behoort juist niet tot de regenwormen (familie Lumbricidae), maar tot de familie Megascolidae.
Het lichaam van de regenworm kan bestaan uit meer dan honderd segmenten, de gewone regenworm heeft er ongeveer 150. Het aantal segmenten neemt met het ouder worden toe doordat in een speciale groeizone vlak bij het uiteinde nieuwe segmenten ontstaan. Vooraan, na ongeveer het dertigste segment, zit bij volwassen regenwormen een verdikking, het clitellum of zadel genoemd.
De huidlaag wordt de epidermis genoemd, aan de buitenkant zit een eencellige laag van epidermiscellen, waarin enkele kliercellen- en zintuigcellen voorkomen. De epidermis is bedekt met een verharde laag die de cuticula wordt genoemd, deze was-achtige cuticula beschermt de worm tegen uitdroging. Onder de epidermis zit een laag dwarsgestreept spierweefsel, met daaronder een dikke laag glad spierweefsel. Dicht onder de huid van het gehele lichaam van de regenworm zit een laag van huidspieren.
Het lichaam van regenwormen is net als bij alle ringwormen opgebouwd uit segmenten of ringen. De gewone regenworm heeft er 110 tot 180, het aantal varieert enigszins per soort. De segmenten zijn binnenin het lichaam gescheiden door een segmentwand, of septum. Deze segmentwanden worden gepenetreerd door de spijsverteringskolom, de aderen, de zenuwstreng en tevens door de nefridiën. Het segment waarin een nefridium is gelegen bevat het blaasje en het kanaaltje van het uitscheidingsorgaan, maar er is steeds een uitloper aanwezig die uitmondt in het ervoor gelegen segment.
Het voorste segment bevat de mondopening. Vóór het voorste segment is het mondflapje aanwezig, dit wordt het prostomium genoemd wat letterlijk voor (pro) de mondopening (stoma) betekent. Een verouderde naam voor het prostomium is 'acron'.[3] Het prostomium is geen echt segment maar een zogenaamd pseudosegment, het is een uitgroeisel van de monddelen.[4] Binnen de familie van de regenwormen komen soorten voor waarbij het prostomium een lobje van het eerste segment is (epiloob) en er zijn soorten waarbij het prostomium een verbinding heeft met het tweede segment (tanyloob). Dit is rechts afgebeeld in het schema. Een derde groep is zygoloob, hierbij hebben de eerste segmenten geen verbinding met elkaar. Achter het prostomium is het eerste echte segment aanwezig, het peristomium, hierin is de mondopening gelegen.[4]
De segmenten van alle regenwormen hebben achter de voorzijde, vanaf het vierde of vijfde segment, gepaarde openingen aan de buikzijde die de nefridioporiën worden genoemd. Deze poriën zijn de uitgangen van de nefridia, dit zijn uitscheidingskliertjes die een rol in de waterhuishouding en de primaire stofwisseling spelen.
De lichaamssegmenten van regenwormen zijn steeds voorzien van kleine, verharde en borstelachtige uitsteeksels die de setae worden genoemd. Bij wormen wordt voor deze dikke, haarachtige structuren ook wel de naam chaetae gebruikt.[5] Ze bestaan uit chitine en eiwitten. De regenworm heeft altijd vier paar borstels per lichaamssegment, alleen op het eerste en het laatste lichaamssegment ontbreken dergelijke structuren. De borstels zijn bij de gewone regenworm gegroepeerd in rijen; een dubbele rij op de buikzijde, de ventrale borstels en een rij aan iedere lichaamszijde, de laterale borstels.[6] Andere soorten regenwormen hebben een afwijkende configuratie van de borstels, zoals alle rijen aan de onderzijde of de borstelharen zijn meer verspreid over het lichaam.[5]
De borstels van een aantal segmenten zijn duidelijk langer dan die van de meeste andere segmenten, deze chaetae worden gebruikt om het lichaam aan een partner te ankeren tijdens de paring. Deze chaetae worden daarom wel de geslachtschaetae genoemd, het aantal en de positie op de lichaamssegmenten van deze borstels zijn per soort verschillend.[4]
De bewegingen van de regenworm worden veroorzaakt door spiersamentrekkingen waardoor het lichaam wordt verplaatst. De stand van de borstels speelt een rol betreffende de richting van de verplaatsing van het lichaam. De borstels geven de regenwormen grip bij het graven, ze staan meestal achterwaarts gericht. Als men een regenworm van voor naar achteren tussen de vingers laat glijden geven de borstels weinig weerstand maar als men dit van achteren naar voren probeert wordt de aanwezigheid van de borstels duidelijk merkbaar. De borstels kunnen wat worden uitgestoken zodat de grip wordt vergroot. Doordat de borstels verhinderen dat het lichaam naar achteren wordt verplaatst kruipt de worm vanzelf voorwaarts. Als de worm zich achterwaarts wil bewegen, bijvoorbeeld om zich terug te trekken in het hol, wordt de stand van de borstels veranderd waarbij ze naar voren wijzen en wordt voorkomen dat het lichaam zich naar voren verplaatst. De manier waarop regenwormen zich voortbewegen wordt wel peristalsis genoemd.
Het zenuwstelsel bestaat uit een zenuwstreng die door het gehele lichaam loopt en in ieder segment vertakkingen heeft. Op de grens van twee segmenten is een grote vertakking aanwezig en in het midden van ieder segment ontspruit een tweede vertakking.
De regenworm heeft twee soorten spieren onder de huid; de spieren die om de segmenten liggen worden de ringspieren genoemd en de spieren die zich in de lengte bevinden zijn de lengtespieren.[7] Bij het graven of kruipen worden de borstels naar achteren gericht zodat het lichaam wordt verankerd. De lengtespieren ontspannen zich terwijl de ringspieren samentrekken, hierdoor wordt het lichaam zowel langer als dunner.
Het voedsel van de regenworm wordt opgenomen door de mond, waarna het langs verschillende organen wordt gevoerd en het afval wordt aan de achterzijde weer afgescheiden. De kop van de regenworm zit altijd aan de zijde van het clitellum, de kop is verder nauwelijks te onderscheiden van de achterzijde. Onder een microscoop wordt de structuur van de kop beter zichtbaar, de worm heeft een soort lapje voor de mond waarmee voedsel kan worden vastgegrepen. Verharde structuren zoals tanden of kaken ontbreken echter. Een regenworm beschikt niet over ogen of oren maar kan waarschijnlijk wel bepaalde chemische verbindingen waarnemen. Zo weet de worm of het voedsel dat wordt gegeten wel geschikt is, er is een voorkeur voor bladeren ten opzichte van takjes.
De 'lip' van de worm wordt het prostomium genoemd en hangt aan het kopeinde over de mond. De mondopening begint net na het eerste segment en gaat door tot in de darm, die door het gehele lichaam van de regenworm loopt. Het begin van de darm bestaat uit een spierloze keel (farynx), die overgaat in een slokdarm (oesofagus). Deze slokdarm bestaat achtereenvolgend uit kalkzakjes, een spierloze 'krop' en gespierde magen. Hierin wordt het plantaardige voedsel met behulp van zandkorreltjes gelijkmatig vermalen (op dezelfde wijze als bij kippen maar die doen het met behulp van steentjes). Hierna zit de lange middendarm, die over de hele lengte op de rugzijde een uitstulping heeft, die de tyflosole wordt genoemd. Deze structuur geeft de middendarm een inwendig grotere oppervlakte. Aan de buitenzijde van de tyflosole zijn de chloragogeencellen aanwezig, die geelbruine van kleur zijn en een functie als uitscheidingsorgaan hebben.[6]
In elk segment met uitzondering van de eerste drie tot vijf en het laatste segment, zijn aan weerszijden van de darm de uitscheidingsorganen gelegen. Deze worden de nefridia genoemd en ze bestaan uit een lang, in lussen gevouwen kanaal met aan het eind een blaasje. Een trechter met wimpers die in het voorliggende segment is gelegen vangt de uitscheidingsstoffen op waarna ze in het kanaaltje terechtkomen. Elk uitscheidingsorgaantje is voor de zuurstoftoevoer omgeven door bloedvaten. In de nefridia vinden veel omzettingsprocessen plaats waarbij energie benodigd is, wat de bloedvaatjes verklaart. In de nefridia worden stoffen als urinezuur, zouten en ammoniak afgevoerd en in de orgaantjes wordt water teruggewonnen uit de afvalstoffen. De nefridia zijn vergelijkbaar met de nieren van hogere gewervelden; ze hebben geen directe interactie met de vertering van het voedsel maar bewerken de uit het voedsel verkregen verbindingen om ze geschikt te maken voor de lichaamsprocessen van de worm. Het uitscheidingsorgaan van het voedsel wordt de anus genoemd en is aan de achterzijde gelegen aan het laatste segment, dit wordt wel het pygidium genoemd. In de uitwerpselen worden vaak nematoden gevonden, kleine wormen, zoals soorten uit het geslacht Rhabditis.
Tussen de darm en de huidspierlaag zit in elk segment de secundaire lichaamsholte, het coeloom. De secundaire lichaamsholten van de aangrenzende segmenten worden door dunne dwarswanden (dissepimente) van elkaar gescheiden. De secundaire lichaamsholten zijn met een vloeistof gevuld en staan onder druk, waardoor er een hydrostatisch skelet gevormd wordt dat de regenworm de nodige stevigheid geeft.
Regenwormen hebben een hoog ontwikkeld zenuwstelsel, net als andere ongewervelde dieren hebben ze geen hersenen maar een aantal knooppunten waar de zenuwen samenkomen. Een dergelijk knooppunt wordt een ganglion genoemd, meervoud ganglia. De regenworm heeft dergelijke knooppunten vooraan de kop, deze worden de hersenganglia genoemd. De ganglia liggen in het derde segment vlak voor het begin van de keel (farynx) aan de rugzijde (dorsaal) van de darm. Van daaruit gaan talrijke zenuwen naar de bovenlip of prostomium. Het prostomium is een belangrijk lichaamsdeel omdat het dient als een tastzintuig bij het zoeken naar voedsel. De bovenlip dient tevens als grijporgaan bij het opnemen van voedsel. De hersenganglia gaan over in een hoofdzenuwstreng die aan de buikzijde doorloopt naar de achterzijde van het lichaam. De centrale zenuwstreng heeft de vorm van een paarsgewijs opgezet touwladderzenuwsysteem. In ieder segment zijn vertakkingen naar de huidspierlaag om de voortbewegingsspieren en de borstelspieren aan te sturen. Deze deelganglia worden ook wel de segmentaire ganglia genoemd. Ze zijn zeer klein en bij een dissectie met het blote oog nauwelijks te zien.
Regenwormen hebben nooit ogen, maar zijn wel gevoelig voor licht. Ze zijn negatief fototactisch; dit wil zeggen dat ze licht proberen te vermijden. Zowel aan de voorzijde als de achterzijde zijn lichtgevoelige zintuigcellen gelegen. Deze cellen worden ook wel de fotoreceptors genoemd. Ook is de regenworm gevoelig voor trillingen, dit dient om gravende dieren die wormen eten op te merken zodat de regenworm snel naar boven kan vluchten.
De regenworm heeft geen speciale ademhalingsorganen, maar wel een sterk vertakt gesloten bloedvatensysteem, waarmee door de huid zuurstof opgenomen wordt en uit de darm opgenomen voedingsstoffen worden getransporteerd. Regenwormen hebben twee bloedvaten die door het gehele lichaam lopen, aan de bovenzijde is het rugvat gelegen dat het bloed van achter naar voren stuwt. Aan de buikzijde is het buikvat gelegen waardoor het bloed van voren naar achteren stroomt. Deze bloedvaten worden in ieder segment met elkaar verbonden door tussenkanaaltjes, de ringvaten. Het rugvat pompt door samentrekking van bepaalde ringvaten het bloed naar voren in de harten. De ringvaten zijn op de plaats van het spijsverteringsorgaan sterk vergroot en hebben een pompfunctie. Ze worden wel laterale harten of pseudoharten genoemd. Het aantal harten varieert per regenworm, de gewone regenworm (Lumbricus terrestris) heeft vijf paar en dus tien harten in totaal.
Om het lichaam van zuurstof te voorzien, hebben regenwormen net als gewervelde dieren hemoglobine als zuurstofdragende stof. De hemoglobine is een pigment dat zuurstof makkelijk bindt en weer kan afgeven, het is bij gewervelden in speciale cellen geborgen, de rode bloedcellen. Bij de regenwormen komen geen rode bloedcellen voor, de zuurstof wordt met behulp van eiwitten in het bloed opgelost. De hemoglobine is gebonden aan een eiwitcomplex dat erythrocruorin wordt genoemd.[8] Deze eiwitten zijn heel complex en kunnen erg groot zijn, het geheel van eiwitten kan uit 36 verschillende delen bestaan en dient als een soort raamwerk om de hemoglobinemoleculen te binden. Omdat de moleculen altijd in twaalf rijen van 12 zijn verdeeld, bevat het eiwitcomplex altijd 144 hemoglobinemoleculen.[4]
Omdat regenwormen soms in zuurstofarme omstandigheden belanden, zoals bij langdurige overstromingen van het land, heeft het bloed een relatief groot zuurstofbindend vermogen.
De bloedvloeistof bevat ook cellen die tot het afweersysteem behoren, de zogenaamde fagocyten. Deze breken lichaamsvreemde en ongewenste deeltjes af, zoals afvalstoffen van cellen en micro-organismen.
Regenwormen zijn hermafrodiet, dit betekent dat ze zowel mannelijke als vrouwelijke geslachtsorganen bezitten. De regenworm heeft een duidelijke verdikking aan de voorzijde van het lichaam, die vaak lichter van kleur is ten opzichte van de rest van het lijf. Deze band wordt het zadel genoemd of met een wetenschappelijker woord het clitellum. Het clitellum is bij de gewone regenworm altijd gelegen tussen het 32e en het 37e lichaamssegment, gezien vanaf de kop.[4] Bij andere soorten is de positie van het clitellum verschillend, zodat ze hieraan te onderscheiden zijn.
Vaak wordt gedacht dat het opvallende, verdikte zadel of clitellum de geslachtsorganen bevat maar dit is niet juist. Het clitellum is een conglomeratie van slijmproducerende cellen en speelt weliswaar een rol bij de vorming van een cocon en de afzet van de eitjes maar is zelf geen onderdeel van het geslachtsapparaat. Het slijm dat wordt afgescheiden door het clitellum dient als 'reageerbuis' waarin de eitjes en het sperma samenkomen en droogt later in tot een cocon dat de eieren beschermt tegen uitdroging.
De inwendige testis en een uitwendige opening vormen het mannelijk orgaan. Ovaria, eileiders en een uitwendige opening vormen het vrouwelijke orgaan. Daarnaast hebben ze spermazakjes, waarin het sperma van de partner wordt opgevangen. De dauwpier copuleert boven de grond, andere soorten in de grond.
Regenwormen worden op basis van hun graafgedrag en levenswijze ingedeeld in drie ecologische groepen. De epigeïsche soorten leven op de bodem in de strooisellaag. Ze blijven klein en graven geen gangen, het lichaam is meestal voorzien van een pigmentatie. De epigeïsche soorten zorgen voor een verkleining van bladafval op de bodem. Een voorbeeld is de soort Lumbricus rubellus.[9] De endogeïsche soorten leven in de toplaag van de bodem. Ze graven tunnels die vaak horizontaal zijn en net onder de oppervlakte liggen. De regenwormen die tot deze groep behoren worden middelgroot en hebben geen pigmentatie, zijn vaak bleek van kleur. Endogeïsche wormen breken bladafval af en zorgen voor beluchting van de toplaag. Een voorbeeld is de soort Allolobophora chlorotica. De derde groep zijn de anekische soorten, dit zijn wormen die diepe, verticale tunnels graven. De anekische soorten hebben pigmentatie en worden het grootst. Door de diepe tunnels wordt de bodem beter belucht en kan het water beter worden afgevoerd. Een voorbeeld is de gewone regenworm (Lumbricus terrestris).[9]
Regenwormpopulaties zijn afhankelijk van zowel de fysische als de chemische eigenschappen van de grond, zoals bodemtemperatuur, vochtigheid, zuurgraad (pH), zouten, beluchting en textuur. Daarnaast moet er voldoende voedsel aanwezig zijn en moet de soort zich kunnen voortplanten en verspreiden. Om dit te bereiken moet er regelmatig organisch materiaal aan de grond worden toegevoegd door dit in de grond te brengen of boven op de grond te strooien.
De regenworm speelt een zeer belangrijke rol in het verbeteren van de bodemstructuur. De regenworm graaft lange tunneltjes waarin atmosferische lucht kan doordringen, zodat de bodem wordt belucht. Dit heeft tot gevolg dat aerobe bacteriën dieper in de bodem kunnen leven; dergelijke bacteriën versnellen de afbraak van organische stoffen nog eens. Doordat de regenwormen in hun gangenstelsels bewegen, wordt de lucht verplaatst in de bodem, wat de beluchting ten goede komt. Het gebruik van zware landbouwmachines leidt ertoe dat de lucht uit de bodem wordt geperst, maar door het werk van regenwormen wordt de grond geschikter voor planten om te groeien. De tunnels van regenwormen worden door planten gewaardeerd: ze kunnen hier makkelijk hun wortels in laten doordringen.[10]
Daarnaast wordt tevens de waterhuishouding van de grond beter als er regenwormen in voorkomen. Door de lange tunnels kan de bodem meer water opnemen, afvoeren en vasthouden, al naargelang de omstandigheden. Een andere eigenschap van de regenworm die de bodemgesteldheid ten goede komt is het eten van dode, afgevallen plantendelen. De regenworm zet deze om in mineralen die erg belangrijk zijn voor planten. Hierbij wordt het organische materiaal gedeeltelijk verteerd en de overgebleven aarde goed gemengd weer afgegeven. Regenwormen werden door Charles Darwin beschreven als een 'natuurlijke ploeg', hij schatte dat om de zoveel jaar de volledige oppervlaktelaag van wereld de maag van een aardworm passeert.
De regenworm speelt met name zo'n grote rol in de ecologie van de bodem omdat de soort in enorme aantallen voorkomt. In humusarme grond kunnen 62 wormen per vierkante meter voorkomen en in humusrijke grond zelfs meer dan 432 wormen per m². Volgens een schatting vormen regenwormen ongeveer de helft van de biomassa in de bodem. Op een oppervlakte van 1000 vierkante meter komen gemiddeld ongeveer 125 kilo regenwormen voor, in vruchtbare grond kan dit oplopen tot 3 ton aan regenwormen, al betreft het hier niet alleen de gewone regenworm maar alle soorten aardwormen.[7] In akkers waar veel wormen aanwezig zijn, is de opbrengst aanmerkelijk hoger dan akkers waarin geen wormen leven. Bij het wegen van de uitwerpselen van de wormen op een oppervlak van een vierkante meter gedurende één jaar bleek dat de dieren 4,4 tot 8 kilo produceren, wat neerkomt op een jaarlijkse verwerking van 44 tot 80 ton aarde per hectare.[3]
Regenwormen planten zich geslachtelijk voort waarbij eieren worden geproduceerd die worden afgezet in een slijmachtige massa die uiteindelijk indroogt tot een eicocon. Regenwormen zijn tweeslachtig (hermafrodiet), maar een regenworm kan niet de eigen eitjes bevruchten. Elke regenworm heeft zowel een mannelijk als een vrouwelijk geslachtsorgaan. Een andere bijzonderheid is het feit dat de wormen wel een echte paring kennen, maar de eieren worden toch buiten het lichaam bevrucht tijdens de vorming van de cocon.
Regenwormen kunnen zich in principe het gehele jaar door voortplanten. In veel streken zijn de winters te streng of de zomers te droog om actief te zijn en hier planten de regenwormen zich voornamelijk in de lente en de herfst voort, de rest van het jaar moeten ze zich verschuilen.
Van de meeste andere soorten regenwormen is bekend dat ze ondergronds de paring uitvoeren maar de gewone regenworm is een van de weinige soorten die bovengronds paart. Onder vochtige omstandigheden kruipen de dieren naar boven en komen uit hun gang op zoek naar een partner. Omdat ze gevoelig zijn voor uitdroging gebeurt dit meestal in de ochtend of avond of overdag na een regenbui. Andere regenwormen leven permanent aan de oppervlakte en paren onder bladafval of ondergronds in een gang.
Tijdens de paring positioneren de wormen zich zo dat de dieren met de lichamen tegen elkaar aan bevinden waarbij de voorzijden tegenovergesteld zijn. Tijdens de paring worden zaadcellen afgescheiden aan de partner. De huidborstels in de omgeving van het clitellum (het zadel) haken zich in de huid van de partner aan de voorzijde van het clitellum. Vervolgens wordt er slijm gevormd door klieren bij de zadels, de slijmlaag vormt uiteindelijk een soort 'wikkel' rond de mannelijke geslachtsorganen van de wormen. Het doel van deze slijmband is het voorkomen dat het zaad zich mengt en de worm alleen het zaad van de partner binnenkrijgt. De paring van de regenworm duurt ongeveer drie tot vier uur.[11]
De mannelijke geslachtsorganen van de regenworm bevinden zich inwendig en hebben een verbinding met een opening aan de buikzijde, de zogenaamde geslachtsporie. De zaadcellen die worden afgescheiden komen terecht in een groef die naar achteren loopt. Doordat deze groef tijdens de paring wordt omgeven door slijm wordt de groef afgesloten en vormt een kanaaltje waardoor het zaad naar de zaadblaasjes van de partner vloeit. Deze zaadblaasjes worden wel spermatotheca genoemd, in deze flesvormige lichaamsholtes worden de zaadcellen levend bewaard.[7] Regenwormen bevruchten elkaar dus niet tijdens de paring maar wisselen alleen zaadcellen uit, de eigenlijke bevruchting vindt later plaats, buiten het lichaam.[3]
Het sperma blijft in de spermatotheca tot de eicellen rijp zijn. Een regenworm kan door deze spermavoorraad na een paring gedurende enkele maanden eieren produceren zonder dat een volgende paring nodig is.[11] Vaak zijn bij een tweede paring nog steeds zaadcellen van een vorige partner aanwezig in de spermatotheek, zodat een soort concurrentie ontstaat tussen de spermacellen.[4]
Als de eieren voldoende zijn ontwikkeld vindt de uiteindelijk de bevruchting plaats. Hierbij wordt opnieuw een slijmlaag rond het clitellum gevormd, dat als een gordel om de worm zit. De slijmband wordt gevormd op de segmenten 8 tot en met 38, maar is het dikst rond het clitellum.[4] In het slijm zijn eiwitten aanwezig welke dienen om de zich ontwikkelende embryo's van voedsel te voorzien. Het slijm bevat tevens een eiwit dat dient als signaalstof. Zodra de slijmband is gevormd wordt deze door de worm achterwaarts verlaten, waardoor de slijmband zich dus naar de voorzijde van het lichaam verplaatst. Zodra de slijmband over de vrouwelijke geslachtopening bevindt, worden de eieren afgezet in het slijm. De vrouwelijke geslachtsopening wordt de gonoporie genoemd en bevindt zich bij de gewone regenworm op het veertiende lichaamssegment.[4]
Nadat de slijmkoker van (nog onbevruchte) eieren is voorzien wordt deze verder afgestroopt en komt even later boven de spermatothecae te liggen. De achterste is bij de gewone regenworm gelegen tussen de segmenten 10 en 11 en de voorste is gelegen tussen de segmenten 9 en 10. Zodra de slijmkoker zich boven de openingen bevindt trekken de blaasjes samen waardoor het sperma vrijkomt en zo de eieren bevrucht.
Als de worm de slijmkoker volledig van het lichaam heeft gestroopt, droogt het uit en vormt een verharde cocon. De cocon heeft ongeveer de grootte van een erwt en heeft vaak een karakteristieke citroenvorm. De cocon heeft een gele tot roodbruine kleur. Hoewel de cocon dus meerdere levensvatbare eieren bevat, kruipt uit de meeste cocons slechts één jonge worm. Bij andere regenwormen zoals Eisenia fetida zitten twee tot vijf eieren in een cocon.[3]
De gewone regenworm kruipt na één tot vijf maanden uit de cocon, afhankelijk van de omstandigheden. Een half jaar tot anderhalf jaar later is de worm geslachtsrijp en kan zich voortplanten.[11] De levensduur van de gewone regenworm in het wild is enkele jaren, maar weinig exemplaren leven lang genoeg om de maximale lengte van 30 centimeter te bereiken. Uit waarnemingen van de worm in gevangenschap blijkt dat een leeftijd van zes jaar kan worden bereikt. Bij andere soorten regenwormen wijkt de ontwikkelingsduur wat af, kleinere soorten regenwormen zijn vaak eerder volwassen maar hebben een lagere levensverwachting. Van erg grote regenwormen zoals de soort Allolobophora longa is bekend dat een leeftijd van meer dan tien jaar kan worden bereikt.
Regenwormen hebben een tamelijk groot herstellingsvermogen of regeneratie, dit is het verschijnsel dat een dier verloren gegane lichaamsdelen kan vervangen. De regenworm kan een deel van het lichaam verliezen door roofdieren die er een stuk af bijten, maar ook door bijvoorbeeld mechanische landbouwtechnieken van de mens worden veel exemplaren in tweeën gehakt. Van de regenworm wordt wel beweerd dat een in twee delen gesneden worm zich tot twee 'nieuwe' wormen ontwikkelt, maar dit is niet juist. In de praktijk gaat de regenworm meestal toch dood bij een lichaamsbreuk, omdat het afgebroken deel te groot is om voor een van beide delen nog levensvatbaar te zijn. Bovendien is vaak ook de inwendige schade groot bij een dergelijk voorval, bijvoorbeeld als een worm uit elkaar wordt getrokken door een vogel. Alleen als er een breuk ontstaat door een niet-traumatische afscheiding, zoals een snede, kan de regenworm het overleven mits het afgenomen deel niet te groot is.
Soorten met een rood gepigmenteerde huid bezitten dit vermogen niet.[12] Regeneratie vindt bijna uitsluitend plaats aan verwondingen aan het achterlijf. Doordat zich ter plaatse van de verwonding de bloedvaten samentrekken verstoppen rode bloedcellen de kapotte vaten, hierna kan weer herstel optreden. Het duurt na de regeneratie nog lang (2-3 maanden) voordat het weer aangegroeide deel de oorspronkelijke kleur terug heeft. Aan de voorkant kunnen tot maximaal vier segmenten verwijderd worden, die daarna gewoon weer opnieuw gevormd worden. Worden er meer verwijderd, maximaal 15, dan kunnen ze niet allemaal weer aangroeien. Als er nog meer segmenten worden verwijderd gaat de regenworm dood. Aan het achtereinde kunnen veel meer segmenten verwijderd worden. Van de inwendige organen is bekend dat het zenuwstelsel kan regenereren, bij de primaire geslachtsorganen is dat niet het geval. Regeneratie vindt alleen plaats bij gunstige omstandigheden, vooral tijdens de warme zomermaanden, maar lang niet altijd.[12]
Regenwormen zijn afvaleters die leven van dood organisch plantaardig materiaal. Plantenresten en bladeren die op de bodem liggen worden in een woongang getrokken en gegeten. De regenworm is niet aan te merken als schadelijk.
Bij het graven van de gangen wordt het meeste materiaal opgenomen maar de grond bevat maar een deel van het benodigde voedsel. Plantendelen die in het hol zijn getrokken worden voorverteerd door afscheidingen van de mond en bodembacteriën voordat ze in het spijsverteringsstelsel terechtkomen. Uit een experiment gedaan door Charles Darwin bleek dat als driehoekige voedseldelen worden aangeboden, deze meestal met de punt naar beneden een gang in worden getrokken. Regenwormen zouden daarom volgens Darwin enige vorm van intelligentie kunnen worden toegedicht.[13] Later werd duidelijk dat de regenwormen simpelweg net zo lang aan een voedseldeeltje trokken tot de minste weerstand was bereikt. De regenworm vindt een punt aan een voedseldeel dus door veel proberen en niet door intelligentie.
Regenwormen vermijden het eten van de uitwerpselen van andere exemplaren. Ze eten soms wel uitwerpselen van andere dieren, omdat deze veel verteerbare stoffen bevatten. Ook de weke delen van dode dieren worden gegeten, ook van andere regenwormen.
Regenwormen staan aan de basis van vele voedselketens en dienen als voedsel voor veel vogelsoorten, zoals de merel, kraai, roodborstje. Ook bij egels, dassen en mollen staan regenwormen op het menu. Verschillende insecten (waaronder kevers), naaktslakken en platwormen hebben regenwormen op het menu staan.
Regenwormen hebben vele vijanden, veel dieren die regenwormen eten hebben zich gespecialiseerd in het opsporen en vangen van de regenworm. De regenworm heeft met name te vrezen van kleine zoogdieren zoals muizen en mollen. Voorbeelden zijn de spitsmuis en de Europese mol, die grote hoeveelheden regenwormen eten. Ook verschillende vogels jagen intensief op de regenworm, een voorbeeld is de merel. Zelfs van grote roofvogels zoals de buizerd is bekend dat zij in voedselarme perioden veel regenwormen eten.[14] Als een regenworm door een vijand in twee delen wordt gehakt kan het verloren gegane deel worden vervangen, zie ook onder regeneratie.
Een nieuwe bedreiging is de invoer van exoten zoals de Nieuw-Zeelandse platworm Arthurdendyus triangulatus, die van regenwormen leeft en buiten zijn natuurlijke verspreidingsgebied geen vijanden kent. De platworm werd in Engeland in 1960 per ongeluk met besmette grond ingevoerd en is nu een plaag. Regenwormen worden geparasiteerd door algen (protozoa), Platyhelminthes en aaltjes. Ze worden gevonden in het bloed, spieren, maag en darm van de volwassen regenworm en in de cocons.
Kortschildkevers
Loopkevers
Amfibieën
Platwormen
Naast gewervelde dieren zijn er ook vele ongewervelden die op regenwormen jagen. Een beruchte vijand zijn loopkevers zoals schallebijters en met name hun roofzuchtige larven. Ook duizendpoten zoals de tuinduizendpoot (Geophilus longicornis) eten regenwormen.[3] Ook sommige bloedzuigers - die net als de regenworm tot de ringwormen behoren- jagen op regenwormen waarbij de worm in één keer wordt verzwolgen. Sommige soorten huisvliegen planten zich voort door eieren op regenwormen te leggen.
Naast ziekten en predatie worden regenwormen bedreigd door het gebruik van kunstmest en gewasbeschermingsmiddelen. Regenwormen kunnen gifstoffen in hun lichaam opslaan waardoor dieren die wormen als voedsel gebruiken vergiftigd kunnen worden.
Regenwormen danken hun naam aan het feit dat ze vooral te zien zijn als het regent en alleen dan over het bodemoppervlak kruipen. Regenwormen kunnen een regenbui waarnemen door de trillingen in de bodem die veroorzaakt worden door de vallende regendruppels. Een bekend misverstand over regenwormen is dat ze proberen te ontsnappen aan de regen omdat ze kunnen verdrinken als hun holletje volloopt. De regenworm leeft vaak in waterige omstandigheden zoals onder de grondwaterspiegel. Ze nemen zuurstof op door de dunne huid, het ademhalingsapparaat werkt echter ook onder water. Aangezien regenwater rijk is aan zuurstof hebben regenwormen niet veel te vrezen van een bui. Als echter zuurstofarm grondwater omhoog komt kan een regenworm verdrinken en zal naar de oppervlakte kruipen.[4]
Regenwormen komen bovengronds tijdens een bui omdat ze regen verwarren met de trillingen van een vijand, zoals een gravende mol. Deze trillingen kunnen worden nagebootst door een stok in de grond te steken en deze te laten trillen. De regenwormen zullen dan massaal naar boven kruipen, ongeacht de weersomstandigheden. De regenworm kan zich zonder gevaar over land verplaatsen en ook de paring vindt bovengronds plaats onder natte weersomstandigheden. Regenwormen zijn veel gevoeliger voor uitdroging dan voor verdrinking. Een belangrijke vijand van de regenworm is het weer. Als het te droog is moeten de wormen dieper in de grond schuilen om niet uit te drogen. De worm manoeuvreert zijn lichaam in een balvorm en vormt een laagje uitwerpselen om het lijf zodat de worm in een soort cocon zit. Hier schuilt de regenworm tot betere tijden aanbreken. In de winter kruipen de wormen dieper in de bodem om aan bevriezing te ontkomen.
De meeste soorten regenwormen produceren calcitische kristallijne korrels.[15] Alle korrels producerende soorten bezitten daarvoor twee klieren dicht bij de kop waarin microkristallijne korrels worden aangemaakt met een grootte van 1–10 micrometer. Deze korrels worden vaak direct in de darm geloosd maar bij veel soorten worden zij in een ander deel van dezelfde klieren tot een aggregaat van korrels van maximaal ongeveer 2,5 millimeter grootte omgezet.[16] De korrels worden uiteindelijk uitgescheiden: uitwerpselen die boven de grond worden afgezet kunnen er honderden van bevatten. Het is onbekend waartoe de korrels dienen maar er is wel een relatie gevonden met de hoeveelheid humus in de bodem: als er veel humus aanwezig is, dan worden meer korrels geproduceerd. Mogelijk dienen ze ter neutralisatie van humuszuren.
De korrels kunnen goed fossiliseren en worden dan ook veel in Kwartaire en Tertiaire sedimenten aangetroffen. Sommige afzettingen kunnen er enorme hoeveelheden van bevatten: in één geval werden 30.000 exemplaren per liter sediment geteld.[17] Door paleomalacologen werden de korrels vroeger foutief aangezien als de rudimentaire schelp van de naaktslakkenfamilie Arionidae. In een fossiele associatie van land- en zoetwatermollusken worden de regenwormkorrels namelijk vaak tezamen met de fossiele schelpen aangetroffen en van Arionidae is bekend dat inwendig onder het 'schild' soms korrelige structuren van kalk aanwezig zijn. Deze resten zijn echter weinig massief en fossiliseren niet. In de vijftiger jaren van de twintigste eeuw werd pas duidelijk dat dit inderdaad geen resten van naaktslakken maar van regenwormen waren.[18] Deze publicatie is echter geruime tijd onopgemerkt gebleven waaraan in 1964 een eind kwam.[19]
Regenwormen spelen een grote rol in de ecologie van de bodem en staan bekend als een van de belangrijkste natuurlijke bodemverbeteraars. Ze spelen vooral een belangrijke biologische rol als afbrekers van dode resten. De regenworm is zeer belangrijk voor de afbraak van organisch materiaal (compostering).[20] Hierbij wordt dood materiaal omgezet in humus en een humusrijke grond is belangrijk voor een goede plantengroei. Uitwerpselen van wormen bevatten soms 40% meer humus dan de bovenste grondlaag, waarin de regenworm leeft. Regenwormen trekken 's nachts bladeren, mest en ander dood organisch materiaal de grond in om later als voedsel te dienen en voor bekleding van het nest. De regenworm kan het voorstuk knopvormig laten opzwellen, waardoor de mond door een zuignap omgeven wordt. Met behulp van de keel zuigt de regenworm zich vast en trekt achteruitkruipend het blad de grond in. Uitscheidingen van de keelklieren bevorderen de vertering.
Het nut als opruimer werd wat genuanceerd door Duits onderzoek van de gewone regenworm; wormen zouden ook levende zaden en zaailingen eten. Dit vormt een potentieel gevaar voor planten uit gebieden waar de soort als exoot voorkomt; voor de meeste gebieden zijn ze echter wel nuttig.[21] Ook chemisch gezien veranderen de regenwormen de bodem; tegelijk met organisch materiaal eet de regenworm ook gronddeeltjes op. In de krop of farynx worden de gronddeeltjes door de daar aanwezige zandkorrels vermalen tot een fijne pasta, waardoor de mineralen voor de plant beter beschikbaar worden. Onderzoek in de VS heeft uitgewezen dat uitwerpselen van de regenworm tot vijf maal meer stikstof, zeven maal meer fosfaat en elf maal meer kalium bevatten dan de omringende grond. Geschat wordt dat regenwormen op deze manier in Nederlandse weidegronden 85 tot 170 kg organisch stikstof per hectare per jaar omzetten in mineraal stikstof (voor de plant opneembaar).[22]
Als regenwormen in grote aantallen in kleigrond leven, kunnen ze een nadelig effect hebben op de bodem van akkerbouwgebieden. De slijmsporen van de regenwormen verharden (verkitten) waardoor grote klompen klei ontstaan die moeilijk van de te oogsten producten gescheiden kunnen worden.[23] Ook wordt het ploegen in dergelijke grond moeilijker. Een mogelijke oplossing is het uitstrooien van brandkalk op het land, dit is een mengsel van calciumoxide en magnesiumoxide, wat de klei losser maakt. Op de regenwormen zelf heeft brandkalk geen effect.[23]
Verschillende soorten worden gebruikt voor het afbreken van organisch afval in o.a. een wormenbak, dit wordt ook wel een vermicultuur genoemd. Het meest geschikt voor een wormenbak is Eisenia fetida, die duidelijk verschilt van de in de grond levende regenwormen.
Regenwormen worden over de gehele wereld verkocht als natuurlijke bodemverbeteraars. In 1980 werden bijvoorbeeld 370 miljoen wormen door de Verenigde Staten gekocht van Canada. Regenwormen worden tevens als aas bij het sportvissen gebruikt. Vooral de soorten Eisenia hortensis en Eisenia fetida zijn hiervoor geschikt. Deze regenwormen worden onder andere ook in België en Nederland gekweekt. Ten slotte zijn regenwormen ook een ideaal voer voor sommige vogels en een aantal in gevangenschap gehouden amfibieën, zoals salamanders.
De regenwormen, maar ook alle andere diepgravende aardwormen, spelen een grotere rol in de cultuur dan men op het eerste gezicht zou denken. Ze zijn niet alleen belangrijke bodemverbeteraars die de oogst kunnen vergroten maar ondergraven ook op de bodem liggende voorwerpen zodat deze in de bodem terechtkomen. Het kan dan gaan om stenen, die worden ondergraven en langzaam maar zeker in de bodem zakken, ook allerlei menselijke voorwerpen zoals sieraden en munten zijn dankzij de activiteit van regenwormen in de bodem terechtgekomen en zijn zo bewaard gebleven.[24]
De Nederlandse naam 'regenwormen' slaat op het feit dat de wormen voornamelijk na een regenbui te zien zijn. In de Nederlandse taal worden alleen de vertegenwoordigers van de familie Lumbricidae als regenwormen beschouwd, ook in de Duitse taal (Regenwürmer) is dit het geval. In de Engelse taal is de term 'regenworm' minder gebruikelijk, hier worden de meeste families, dus niet alleen de Lumbricidae, uit de onderklasse Oligochaeta met aardworm (earthworm) aangeduid. Er zijn ongeveer 670 soorten regenwormen, waarvan er 22 voorkomen in Nederland.[2]
De wetenschappelijke naam van de groep waartoe de regenwormen behoren, Oligochaeta, verwijst naar de rijen kleine borstels op het lichaam. In vergelijking met andere wormen hebben regenwormenachtigen relatief veel minder borstels, oligo betekent 'weinig' en chaeta betekent 'borstels'. Een andere groep van ringwormen zijn de borstelwormen (Polychaeta) en deze wormen hebben juist veel borstels, poly betekent veel. De gewone regenworm heeft als wetenschappelijke naam Lumbricus terrestris en dit betekent letterlijk worm (Lumbricus) van de aarde (terrestris).
Regenwormen worden net als alle andere dieren in verschillende groepen ingedeeld, zoals stam, klasse en familie. De hoogste diergroep waartoe de regenwormen worden gerekend is de stam van de ringwormen (Annelida). De meeste ringwormen leven in zee, maar bepaalde groepen kunnen in zoet water of zelfs op het land leven. Regenwormen worden verder bij respectievelijk de klasse Clitellata of zadeldragers en de onderklasse Oligochaeta of borstelarme ringwormen ingedeeld. De vertegenwoordigers van de Oligochaeta die een segmentering van het lichaam hebben, zoals de regenwormen, worden verder tot de orde Haplotaxida gerekend.
De haplotaxiden zijn een gevarieerde groep van wormen, die zowel in het water als op het land leven. De terrestrische soorten leven altijd in vochtige omgevingen, zoals modder, mest of compost. De regenwormen (familie Lumbricidae) zijn slechts een van de meer dan tien families van de Haplotaxida. Andere bekende families zijn de Naididae, waartoe de beekslingerwormen uit het geslacht Tubifex behoren en de familie Enchytraeidae waartoe alle enchytreën (potwormen of witte wormen) worden gerekend. Soorten uit de familie Haplotaxidae zijn bewoners van grondwater en diepe meren, deze soorten zijn lang, tot dertig centimeter, maar zijn slechts enkele millimeters in doorsnede.
De verschillen tussen de families van de Haplotaxida zijn anatomisch gezien soms verstrekkend. Bepaalde lichaamsdelen komen niet voor of andere zijn juist extreem ontwikkeld. Een voorbeeld zijn de vertegenwoordigers van de familie Criodrilidae, die geen tyflosole hebben en tevens ontbreekt een spiermaag, ze bezitten echter wel een hoorn-achtige spermatofoor wat ongebruikelijk is bij ringwormen.
Andere niet direct aan de regenwormen verwante families lijken uiterlijk juist sterk op regenwormen, zoals de broze slibworm (Lumbricilus variegatus), die behoort tot de familie van de Lumbriculidae. Vertegenwoordigers van deze groep leven in zeer vochtige omstandigheden en zijn eerder aan te merken als waterbewonend dan bodembewonend. De soorten uit deze familie hebben echter geen duidelijk zadel of clitellum.
Regenwormen (Lumbricidae) zijn een familie van ongewervelde dieren die behoren tot de ringwormen (Annelida). Regenwormen behoren tot de Oligochaeta, dit is een groep van wormen die de zee verlaten hebben en in zoetwater maar ook op het land kunnen leven. Regenwormen komen wereldwijd algemeen voor, uitgezonderd in droge klimaten en op Antarctica.
Meestal zijn ze in grote aantallen te vinden, enkele soorten zijn wereldwijd uitgezet om de bodemstructuur te verbeteren. Regenwormen hebben door het graven van tunnels en het afbreken van plantaardig materiaal een grote invloed op de bodemstructuur. Wereldwijd zijn er ongeveer 670 soorten regenwormen bekend, die in lengte variëren van enkele centimeters tot decimeters.
Bij het trillen van de bodem kruipen de regenwormen uit de grond, dit kan door de loopbewegingen van dieren worden veroorzaakt of door menselijk handelen zoals met behulp van een stok of greep. Onder natuurlijke omstandigheden komen regenwormen boven door graafbewegingen van mollen. Regenwormen komen ook bovengronds bij regenachtig weer om te paren, ze zijn verder vooral 's nachts actief. In de winter kruipen ze dieper de grond in en gaan in een soort winterslaap of diapause. Sommige gewervelde dieren zijn sterk afhankelijk van de regenworm omdat ze een belangrijke voedselbron vormen, voorbeelden zijn de mol en de merel.
Meitemarker er en familie av leddormer som lever i jord, og er viktige for jordkvaliteten, fordi de drenerer og lufter jorda.
Ordene «mark» og «makk» blir benyttet endel i dagligtale, for å referere til meitemark.
Meitemarker har ikke øyne, men lysømfindtlige celler (fotoreseptorer) i huden, særlig på den fremre delen av kroppen.
De største meitemarkene i verden finnes i Australia, og skal kunne bli flere meter lange.
Meitemarken lever av organisk materiale, som råtnende planterester. De drar med seg dødt løv ned under bakken, og er dermed med på å tilføre jorden ekstra næringsstoffer. De graver og lager ganger som gjør at det kommer luft ned i jorden. Dette er nyttig for plantene som vokser der.
Meitemarker finnes helst i feit, fin matjord, men de er nesten over alt, selv i ganske sandholdig eller sur skogsjord.
Meitemarken puster med huden. Alle levende celler må være i et fuktig miljø for å fungere. Derfor må meitemarken passe på at huden ikke tørker ut. Nede i jorden er meitemarken beskyttet mot solys, og det er nok fuktighet slik at hudcellene ikke tørker ut.
Når det er kraftig regnvær ser man ofte mange meitemark på overflaten. Grunnen til dette er uvisst, men det kan være at gangene til meitemarken blir fylt med vann slik at marken drukner. Mye kan likevel tyde på at dette hovedsakelig er for å finne maker. Observerer man meitemark som ligger helt eller delvis over bakken på fuktige, våte netter, ser man ofte to og to som henger sammen.
Det er 19 arter i Norge[1]. Den lengste kan bli 30 cm.
De vanligste er:
M. Weis Clausen. Regnorme (dansk). 84. Danmarks Fauna.
Meitemarker er en familie av leddormer som lever i jord, og er viktige for jordkvaliteten, fordi de drenerer og lufter jorda.
Dżdżownicowate, dżdżownice (Lumbricidae) – rodzina skąposzczetów przystosowanych do życia w glebie. Obejmuje ponad 670 gatunków, w tym większość glebowych skąposzczetów. Dżdżownicowate są jednym z ważniejszych czynników wpływających na przewietrzanie i nawożenie gleby. Są hermafrodytami (obojnakami), ale występuje u nich zapłodnienie krzyżowe. Nieliczne rozmnażają się partenogenetycznie.
Pierwszą poważną pracą poświęconą dżdżownicom i ich wpływowi na glebę była książka Karola Darwina The Formation of Vegetable Mould through the Action of Worms.
Występują bardzo licznie w umiarkowanej strefie Holarktyki. W 1 m³ gleby przebywa przeciętnie 800 osobników[1] w różnych stadiach rozwojowych. Niektóre gatunki (np. Allolobophora caliginosa) są kosmopolityczne. Zasięgi występowania gatunków odpornych na mróz sięgają do strefy tundry w Europie i do arktycznej Syberii[2].
Dżdżownice zasiedlają wszystkie typy gleb. Preferują gleby wilgotne i żyzne, bogate w materię organiczną, o odczynie obojętnym lub lekko zasadowym[2]. Częściej występują w glebach gliniastych. W glebach lekkich, piaszczystych, łatwo przesychających, a także w skrajnie ciężkich żyją tylko nieliczne, przystosowane gatunki. Optymalny odczyn gleby mieści się w zakresie pH 5,5–8. Jednakże są gatunki, które licznie występują także w glebach o pH 3,8 (np. lasy). Niewiele dżdżownic toleruje także wartości pH powyżej 8.
Nieliczne gatunki z tej rodziny żyją w przybrzeżnej strefie wód[2].
W Polsce stwierdzono występowanie 32 gatunków dżdżownicowatych[3], wśród których pospolitych i często spotykanych jest zaledwie około 10. W większości środowisk strefy nizinnej kraju dominują przeważnie tylko 2–4 gatunki. Rozmieszczenie dżdżownic w glebie zależy przede wszystkim od cech poszczególnych gatunków, a w pierwszym rzędzie od ich wymagań pokarmowych. Niektóre odżywiają się na powierzchni i żyją w górnej warstwie gleby, dalej będą one nazywane powierzchniowymi. Osobniki tych gatunków mają wyraźne czerwone lub czerwono-fioletowe ubarwienie części grzbietowej oraz spłaszczony tylny koniec ciała. Drugą grupę stanowią gatunki typowo glebowe, które żyją w głębi ziemi, rzadko wychodzą na powierzchnię i nie mają zabarwienia – będą dalej określane jako glebowe albo glebożerne. Pośrednie miejsce zajmuje dżdżownica ziemna (Lumbricus terrestris) pospolicie nazywana rosówką – największy wśród pospolitych gatunków.
Dżdżownice są pierścienicami od małych aż do średnich i dużych rozmiarów. Ich ciało ma wydłużony, obły kształt i zmienne ubarwienie. Osiąga długość od 1 cm do 1 m, a średnica mieści się w przedziale 1–20 mm[2]. Jest złożone z wyraźnie zaznaczających się segmentów, z którego wystaje 8 małych szczecinek ułożonych parami[4]. W części głowowej znajduje się zwój okołoprzełykowy, pełniący funkcję mózgu, oraz otwór gębowy, wyposażony w małe rogowe szczęki. Mają Hydroszkielet.
Dżdżownice chętnie zasiedlają gleby wilgotne, ponieważ ich ciało zawiera ponad 80% wody, a woda jest stale wymieniana z glebą. W ciągu doby około 60% wody jest wydalane przez układ wydalniczy i uzupełniane wraz z pokarmem oraz przez powłoki ciała.
W przypadku nadmiernego przesuszenia gleby, zbyt wysokiej temperatury i odwodnienia wszystkie dżdżownice mogą zapadać w stan odrętwienia. Może ono trwać nawet kilka miesięcy. Często to samo zjawisko jest wywoływane brakiem pożywienia. Niektóre gatunki glebożerne zapadają w odrętwienie, które nie ma związku z pogarszaniem się warunków bytowania i jest u nich zjawiskiem normalnym. Stan ten trwa 1–2 miesięcy, najczęściej w okresie letnim. Dżdżownice zwijają się w charakterystyczny kłębuszek i pozostają w jamce wysłanej śluzem.
Dżdżownice dość dobrze znoszą dłuższe okresy zalania dobrze natlenioną wodą. Wychodzenie ich na powierzchnię w wilgotne, chłodne noce po ciepłym dniu czasami tłumaczone jest brakiem tlenu w glebie. Bardziej prawdopodobną przyczyną jest jednak działanie siarkowodoru wytwarzanego w ciągu dnia przez drobnoustroje glebowe przy niedoborze tlenu. Większość dżdżownic jest bardzo wrażliwa na działanie niskich temperatur i już lekki mróz (-1 lub –2 °C) jest dla nich śmiertelny. Na polach i działkach pozbawionych okrywy bardzo dużo dżdżownic ginie z powodu wiosennych lub jesiennych przymrozków, a nawet temperatur bliskich 0 °C. Zimę spędzają głęboko w niezamarzniętej glebie albo zapadają w stan odrętwienia.
Nieliczne gatunki przystosowały się do niskich temperatur i są zdolne do przeżycia w zamarzniętej glebie[2].
Dżdżownice są saprofagami – odżywiają się związkami organicznymi zawartymi w rozkładających się roślinach i szczątkach zwierząt. Niektóre, żyjące w głębi gleby korzystają z obumierających korzeni i próchnicy glebowej. Dżdżownice chętnie zjadają wprowadzony do gleby obornik, a także odchody zwierząt na pastwiskach.
W dzień dżdżownice ściółkowe i powierzchniowe przebywają w glebie lub w innym ukryciu, natomiast nocą żerują, wciągając do chodników martwą materię organiczną. W poszukiwaniu odpowiedniego pożywienia wykonują kilkumetrowe wędrówki, ale później często wracają do własnej norki. Wybór pokarmu nie jest przypadkowy – dżdżownice wykazują wyraźne upodobania. Decyduje w tym wypadku przede wszystkim zawartość azotu białkowego i łatwo rozkładalnych węglowodanów.
Większość gatunków potrafi rozróżniać pobierany pokarm. Rosówka chętnie zjada liście szczyru, pokrzywy, bzu czarnego, jesionu, wiązu, głogu, czeremchy. Mniej chętnie wybierane są liście dębu, buka i drzew iglastych, a więc roślin o dużej zawartości garbników i żywic. Jako wyjątkowo smakowite traktowane są liście roślin motylkowych ze względu na wysoką zawartość białka. Wraz z resztkami organicznymi dżdżownice zjadają i trawią liczne mikroskopijnej wielkości organizmy rozkładające te resztki. Są to pierwotniaki, glony, bakterie, grzyby glebowe, promieniowce, nicienie i wiele innych. Organizmy te stanowią główne źródło pokarmu dla gatunków kompostowych.
Trochę inaczej kształtują się wymagania pokarmowe dżdżownic glebożernych, żyjących w głębi ziemi i rzadko wychodzących na powierzchnię. W ich jelicie stwierdzono niewielką ilość substancji organicznej, głównie pochodzących z korzeni i bardzo dużo cząstek mineralnych. Dżdżownice te zjadają w ciągu doby ilość gleby równą 20–40% masy ich ciała; tylko niewielka część zostaje przyswojona, reszta jako gruzełkowate odchody – koprolity – trafia na powierzchnię gleby albo pozostaje w wydrążonych chodnikach.
Wpływ dżdżownic można opisać na przykładzie ich inwazji w lasach umiarkowanych i tajdze Ameryki Północnej, które były pozbawione dżdżownic wskutek zlodowacenia 11 tys. lat temu. Rozwinęły się tam ekosystemy z grubą warstwą powoli rozkładającej się ściółki leśnej. Wpływ inwazyjnych dżdżownic na lasy jest zróżnicowany w zależności od gatunków dżdżownic, właściwości gleby i historii użytkowania terenu. W lasach tych dżdżownice redukują grubość warstw organicznych, zwiększają zbitość gleby, wciągają próchnicę i ściółkę leśną w głąb gleby. Większa zbitość gleby zmniejsza wsiąkanie wody i zwiększa spływ powierzchniowy, co przy zmniejszonej grubości ściółki zwiększa erozję. Zmienione właściwości próchnicy i ściółki leśnej to zmienione warunki kiełkowania nasion, co prowadzi do zaniku organizmów dostosowanych do poprzednich warunków[5]. W niektórych lasach inwazja dżdżownic prowadzi do wpłukiwania azotu i fosforu z warstw gleby o największej koncentracji korzeni[6].
Dżdżownice są w większości obojnakami – każdy osobnik ma gamety męskie i żeńskie. Podczas kopulacji dwa osobniki wymieniają między sobą gamety przywierając brzusznymi stronami ciała. Ciała sklejone są śluzem wydzielanym przez siodełko (kilka zgrubiałych pierścieni tworzących się na oskórku), dlatego proces wymiany gamet nazywany jest siodełkowaniem. Gamety przedostają się do siodełka i są przekazywane partnerowi, który na krótko magazynuje je w zbiornikach nasiennych. Po dokonaniu wymiany zwierzęta się rozchodzą.
Po kilku dniach siodełko wytwarza stwardniałą, błoniastą mufkę wypełnioną gęstą cieczą. Mufka, przesuwana ruchami ciała w kierunku głowy, dociera do żeńskiego otworu płciowego, skąd przedostają się do niej dojrzałe jaja. Po dotarciu do zbiorników nasiennych pobierane są plemniki. Po ześlizgnięciu się z ciała zwierzęcia powstaje, zawierający do 20 jaj, kokon – w nim następuje zapłodnienie. Zabezpieczone substancjami bakteriobójczymi kokony mogą długo przebywać w ziemi, nie ulegając rozkładowi. Z zapłodnionych jaj rozwijają się małe dżdżownice. Ich rozwój do stadium dojrzałego trwa od kilkunastu do 40 tygodni – w zależności od gatunku[4].
Nieliczne gatunki dżdżownic rozmnażają się partenogenetycznie[4].
Dżdżownice glebożerne, zwłaszcza kompostowiec różowy (Eisenia fetida) i Lumbricus rubellus, są wykorzystywane do przetwarzania odpadów organicznych, m.in. do tzw. wermikompostowania komunalnych odpadów organicznych. Ten typ kompostowania z udziałem tych bezkręgowców jest popularny m.in. w ośrodkach miejskich, budynkach mieszkalnych oraz miejscach pracy w Kanadzie[7].
Dżdżownice są stosowane jako przynęta wędkarska, a zmrożone i poszatkowane jako pokarm uzupełniający dla ryb akwariowych.
W rodzinie Lumbricidae opisano około 1130 nazwanych gatunków, z czego, w wyniku rewizji taksonomicznej, 670 uznano za formalnie poprawne, a 55 ma niepewną pozycję taksonomiczną. Pozostałe nazwy są wadliwe lub synonimiczne. W 2005 Robert J. Blakemore specjalizujący się w taksonomii skąposzczetów opracował kompletną listę gatunków zaliczanych do rodziny dżdżownicowatych (lista była ostatnio aktualizowana w 2008)[8].
Wyróżniane są dwie podrodziny:
Pierwsza jest reprezentowana przez jeden rodzaj Diporodrilus, który obejmuje kilka gatunków występujących na Korsyce. Do drugiej zaliczono wszystkie pozostałe dżdżownice w kilkudziesięciu rodzajach, m.in.:
Najliczniejsze w gatunki są rodzaje Allolobophora i Dendrobaena.
Dżdżownicowate, dżdżownice (Lumbricidae) – rodzina skąposzczetów przystosowanych do życia w glebie. Obejmuje ponad 670 gatunków, w tym większość glebowych skąposzczetów. Dżdżownicowate są jednym z ważniejszych czynników wpływających na przewietrzanie i nawożenie gleby. Są hermafrodytami (obojnakami), ale występuje u nich zapłodnienie krzyżowe. Nieliczne rozmnażają się partenogenetycznie.
Pierwszą poważną pracą poświęconą dżdżownicom i ich wpływowi na glebę była książka Karola Darwina The Formation of Vegetable Mould through the Action of Worms.
Família de vermes de terra característica das regiões moderadas do Hemisfério Norte e inicialmente originário da zona paleárctica (Europa, África do Norte, Médio Oriente e Ásia).[1] Compõe o essencial da fauna de vermes de terra européia. Introduzida na América do Norte com a chegada dos primeiros europeus, espalhou-se até a compor hoje a quase totalidade das espécies canadenses ao leste das Rochosas. Foi introduzido igualmente nas regiões temperadas do Hemisfério Sul, onde atualmente prospera (Austrália, África do Sul, Patagônia). Esta família compreende várias centena de espécies.
Família de vermes de terra característica das regiões moderadas do Hemisfério Norte e inicialmente originário da zona paleárctica (Europa, África do Norte, Médio Oriente e Ásia). Compõe o essencial da fauna de vermes de terra européia. Introduzida na América do Norte com a chegada dos primeiros europeus, espalhou-se até a compor hoje a quase totalidade das espécies canadenses ao leste das Rochosas. Foi introduzido igualmente nas regiões temperadas do Hemisfério Sul, onde atualmente prospera (Austrália, África do Sul, Patagônia). Esta família compreende várias centena de espécies.
Daggmaskar (Lumbricidae) är en familj i ordningen Haplotaxida. De är tvåkönade (hermafroditer) med fyra par korta borst på varje segment. De för oss vanligaste arterna av daggmask är mycket viktig för jordens ekologi.
Det finns 2000 sorters daggmaskar i världen, varav 14 i Sverige.[källa behövs] Den längsta i Sverige kan bli 30 centimeter lång. Den största daggmasken är den sydafrikanska jättedaggmasken som kan bli upp till 7 meter lång. Daggmaskar bidrar till nedbrytandet av växtmaterial och på så sätt bidrar till hur den lokala jordmånen ser ut. Systematiken är omstridd.
Arter går att identifiera med en bestämningsnyckel utarbetad av Lofs.[1] Det är ett enkelt schema som tar fasta på detaljer i signalementet, eller beteendet som daggmasken har, art för art. De största i Sverige är Lumbricus terrestris och Allolobophora longa som blir upp till 30 cm långa och är ljusröda, men även kompostmaskar som Eisenia foetida som bara blir 10 cm långa och är nötbruna till lila.[källa behövs]
Daggmaskar (Lumbricidae) är en familj i ordningen Haplotaxida. De är tvåkönade (hermafroditer) med fyra par korta borst på varje segment. De för oss vanligaste arterna av daggmask är mycket viktig för jordens ekologi.
Det finns 2000 sorters daggmaskar i världen, varav 14 i Sverige.[källa behövs] Den längsta i Sverige kan bli 30 centimeter lång. Den största daggmasken är den sydafrikanska jättedaggmasken som kan bli upp till 7 meter lång. Daggmaskar bidrar till nedbrytandet av växtmaterial och på så sätt bidrar till hur den lokala jordmånen ser ut. Systematiken är omstridd.
Arter går att identifiera med en bestämningsnyckel utarbetad av Lofs. Det är ett enkelt schema som tar fasta på detaljer i signalementet, eller beteendet som daggmasken har, art för art. De största i Sverige är Lumbricus terrestris och Allolobophora longa som blir upp till 30 cm långa och är ljusröda, men även kompostmaskar som Eisenia foetida som bara blir 10 cm långa och är nötbruna till lila.[källa behövs]
Lumbricidae (лат.) — семейство почвенных малощетинковых червей из подотряда дождевых (Lumbricina). К этому семейству принадлежат лишённые глаз крупные черви (от 10 до 30 см длиной) с толстой кожей и красной кровью. По бокам сегментов расположено по две пары мелких крючковидных щетинок. Роды и виды этого семейства различаются по форме головной лопасти (так называемой верхней губы), по положению пояска и по числу колец. В России встречаются несколько десятков видов из родов Lumbricus, Dendrobaena и Allolobophora. Представители этого семейства — популярная наживка для рыбной ловли.
Lumbricidae живут в почве, в которой выкапывают длинные трубчатые ходы; ночью выходят на поверхность земли; они втаскивают в свои ходы различные органические остатки — частички листьев и других растительных частей. Питаются разлагающимися органическими веществами. Испражнения этих червей, содержащие в себе много измельчённых земляных частиц, откладываются ими на поверхности земли. Этим они способствуют увеличению пахотного слоя земли, в то время как своими норами разрыхляют почву, а вытаскиванием растительных остатков увеличивают содержание органики. На значение дождевых червей в процессе образования почвы одним из первых указал Чарлз Дарвин (1881).
Передвигается червь благодаря попеременному сокращению и расслаблению кольцевых и продольных мышц. При сокращении кольцевых мышц тело червя становится длиннее и тоньше, а при сокращении продольных мышц — короче и толще.
Каждая половозрелая особь обладает женской и мужской половой системой (синхронный гермафродитизм).
Спаривание происходит ночью на поверхности почвы. Черви плотно прилегают один к другому брюшной стороной таким образом, что протоки семенников каждой особи совпадают с семяприёмниками другой. Каждый червь выделяет специализированными сегментами (пояском) слизистую муфту. После этого сперма одной особи поступает в семяприемники другой, а неоплодотворённые яйцеклетки откладываются в слизь. Оплодотворение происходит при сбрасывании муфты через передний конец тела. В дальнейшем она функционирует как яйцевой кокон.
Lumbricidae (лат.) — семейство почвенных малощетинковых червей из подотряда дождевых (Lumbricina). К этому семейству принадлежат лишённые глаз крупные черви (от 10 до 30 см длиной) с толстой кожей и красной кровью. По бокам сегментов расположено по две пары мелких крючковидных щетинок. Роды и виды этого семейства различаются по форме головной лопасти (так называемой верхней губы), по положению пояска и по числу колец. В России встречаются несколько десятков видов из родов Lumbricus, Dendrobaena и Allolobophora. Представители этого семейства — популярная наживка для рыбной ловли.


_taken_with_a_scanning_electron_microscope.jpg)












.jpg)
.jpg)






.JPG)


























