nomes no trilho de navegação
Glomerella cingulata is a fungal plant pathogen, being the name of the sexual stage (teleomorph) while the more commonly referred to asexual stage (anamorph) is called Colletotrichum gloeosporioides. For most of this article the pathogen will be referred to as C. gloeosporioides. This pathogen is a significant problem worldwide, causing anthracnose and fruit rotting diseases on hundreds of economically important hosts.
C. gloeosporioides has an extremely broad host range, causing anthracnose disease on a variety of crops such as cereals and grasses, legumes, fruits, vegetables, perennial crops, and trees. It has been observed as infecting harvested durian of the species Durio graveolens.[1] Some studies suggest that C. gloeosporioides has sub-populations specific to each host.[2] The symptoms can vary from host to host, but tend to manifest as water soaked, sunken spots on fruit that turn necrotic as the disease progresses, and small dark lesions on leaves.__
Using mangoes, one of the most economically important hosts, as an example, fruit symptoms manifest late in the season as infected fruit ripens to maturity. At this point fruit develops large, sunken areas of decay that are dark brown to black in color. Occasional fruit cracking can also occur when linear necrotic lesions develop into deep cracks through the epidermis that can extend down into the pulp of the fruit. Because symptoms remain unseen before ripening, plants that appear healthy upon picking can become quickly riddled with disease in storage or transport. Mango leaves exhibit symptoms as small, angular, brown/black lesions that enlarge as the disease progresses.[2] Again, these symptoms vary from host to host, but mangoes serve as a decent example for the general symptomatology of this pathogen.
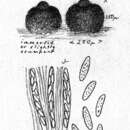
In chestnuts, disease symptoms may also be called blossom end rot. Browning of the chestnut burs at the blossom end may be a first sign in August. At harvest time, blackening of pointed end of the chestnut shell and kernel indicates infection. The extent of blackening can be variable. It can range from a barely visible black tip of the kernel to the whole nut being black. Parts of the nut kernel with no color change remain edible.[3] Regardless of host, C. gloeosporioides produces several signs useful for diagnosis. When observed under a dissecting microscope acervuli can often be spotted if the diseased tissue has recently been under sporulating conditions. These acervuli will appear orange to pinkish in color, due to the masses of conidia being produced on the surface, and will have black, hair-like, setae spiking out in several directions. Under a compound microscope conidia appear ovoid in shape. On certain hosts the teleomorph of this pathogen (G. cingulata) readily produces perithecia full of asci. When cultured on potato dextrose media, this species can appear gray, orange, or pink in color, and will often exhibit concentric rings of growth radiating from the center. C. gloeosporioides can also be identified by PCR if the required resources are available.[4]
The economic impact of C. gloeosporioides varies depending on the host species, but its major impact on some of the more economically important hosts makes the pathogen a huge concern for growers worldwide. For example, in strawberries produced in China, C. gloeosporioides, along with Colletotrichum acutatum, and Colletotrichum fragariae, cause up to 80% of plant loss in strawberry nurseries, and over 40% of yield loss in field.[5] In just these two examples this pathogen is causing millions of dollars in losses.
Recent research suggests that, due to the variability of C. gloeosporioides among hosts, this pathogen should be split into different species or at least different formal species designations. Phylogenetic analysis of C. gloeosporioides isolates sampled from across the broad host range shows enough genetic variation to suggest a need for more specific classification among this species. Work is being done to provide a modern classification system for what is being called the "C. gloeosporioides complex".[6]
The disease cycle is impacted by which form of the pathogen (teleomorph or anamorph) is on the host. This distinction affects how the pathogen overwinters or survives periods without a susceptible host. If the sexual stage (teleomorph) is present, the pathogen sexually reproduces to form ascospores inside of asci, and subsequently packed into perithecia. This provides genetic variation and the convenience of perithecia, which can act as a survival structure. If only the asexual stage (anamorph) is present, the pathogen must survive inside of infected plant tissue or on an alternate host.
Once environmental conditions are met (<95% humidity, 25–28 °C), the ascospores are ejected and infected plant tissue sporulates. Ascospores infect directly, while the infected plant tissue produces acervuli that produce masses of conidia on conidiophores. These conidia are disseminated by rain splash or wind onto new infection courts such as leaves, young fruit, or blossoms.[7] Upon infection, the pathogen continues to produce conidia throughout the season resulting in a polycyclic disease cycle. Once the host plant starts to senesce, the teleomorph form of the disease (G. cingulata) will sexually produce ascospores in perithecia to restart the cycle.
C. gloeosporioides is a hemibiotroph, meaning it lives part way between the biotrophic and saprophytic lifestyles. The pathogen prefers a living host, but once the host tissue dies, or the pathogen finds itself surviving in the soil without a host, it can switch to a saprophytic lifestyle and feed off of dead plant material. Under correct environmental conditions, if a C. gloeosporioides conidium lands upon a susceptible host it will first produce an appressorium. This specialized structure allows the pathogen to penetrate the host cuticle and cell wall through the production of a penetration peg. After penetration, the pathogen produces infection vesicles which invaginate the cell membrane, and drain nutrients from the plant. Later in the pathogen's life cycle, when the host's infected fruit or foliar flesh dies, the pathogen switches to the saprophytic life cycle to feed off of the dead tissue.[8]
This pathogen grows best at 25–30 °C, a humidity>95%, and a pH of 5.8 to 6.5. Pathogenesis can occur at a temperature range as broad as 20–30 °C. Acervuli release spores only when an abundance of moisture is present, so C. gloeosporioides is inactive during the dry season. Direct sunlight, extreme temperatures on either side of the optimum range, and low humidity can all cause spores to become inactive. These three factors can either be extreme enough to cause spore inactivation single-handedly, or work in concert to have the same effect.[7]
Control methods vary depending on the host, but there are some general cultural practices that can be very useful for managing this pathogen. Because C. gloeosporioides spores are spread by rain splash, avoiding overhead irrigation can be helpful. If the operation is small enough the grower might want to consider an overhead covering to avoid rainfall all together. Pruning and thinning out of vegetative material can provide extra airflow, preventing high moisture conditions necessary for disease propagation. Also, to insure a healthy crop to begin with, growers should propagate using only certified, disease-free stock.[9] To prevent post-harvest storage rot, fruit are often subjected to hot water dips or wax coatings. While these methods have been shown to decrease disease incidence, they are not an efficacious solution for the prevention of post-harvest C. gloeosporioides infection.
In terms of chemical control, broad spectrum, protectant fungicides such as chlorothalinil or mancozeb can be applied at the beginning of the growth season to prevent infection. Frequently, once fruit have begun to grow, any further fungicide applications simply mask symptoms until post-harvest. During post-harvest treatment, harvested fruit are often coated with fungicides such as phenols or benzimidazoles to further protect against storage rot, especially on fruit being shipped overseas.[10]
Both cultural and chemical practices need to work together to maintain a safe and practical spraying schedule for a grower. A forecasting system has been built using the humidity and temperature requirements for this pathogen. Using this system a grower can predict what sort of spraying schedule will be most useful for containing C. gloeosporioides.[10]
Glomerella cingulata is a fungal plant pathogen, being the name of the sexual stage (teleomorph) while the more commonly referred to asexual stage (anamorph) is called Colletotrichum gloeosporioides. For most of this article the pathogen will be referred to as C. gloeosporioides. This pathogen is a significant problem worldwide, causing anthracnose and fruit rotting diseases on hundreds of economically important hosts.
Colletotrichum gloeosporioides est une espèce de champignons ascomycètes (syn. Gleosporium olivarum). Il est l'agent de la maladie de l'anthracnose qui touche en particulier les tomates et les olives. Il appartient au genre Colletotrichum dans la famille des Glomerellaceae. Le stade sexué (téléomorphe) correspondant est Glomerella cingulata. Les espèces du genre Colletotrichum sont parmi les principaux agents pathogènes des plantes, dans le monde entier.
Le genre Colletotrichum se caractérise par la formation de structures appelées acervules[1] en forme de disques aplatis, subépidermiques, avec des épines, les conidiophores simples et allongés, avec des conidies hyalines, unicellulaires, généralement en forme de bâtonnets, qui permettent[Quoi ?] aux acervules agrégées par une masse de polysaccharides solubles dans l'eau. Malgré le fait que ces spores ne sont pas des structures de résistance, les mycéliums du champignon peuvent subsister vivants pendant une logue période de temps, dans les semences, les débris culturaux ou en infections latentes dans les fruits.
Le champignon a une croissance rapide en milieu de culture Potato Dextrose Agar (PDA), formant des colonies concentriques de couleur vert-olive à marron, pouvant former ou non des secteurs.
Sous l'appellation d'anthracnose, on trouve une grande variété de fruits tropicaux atteints :
ainsi que d'autres plantes (avocats, passiflores, annone, cacao, lime. Sous des latitudes plus nordiques fraises, pêches, etc. Des légumes sont touchés comme les piments et les tomates. Les pertes économiques sont importantes[2].
Le champignon est l'agent de la Lèpre de l'Olive. La maladie affecte les feuilles et les olives. Le champignon pénètre dans la plante par les ouvertures naturelles (lenticelles, stomates) ou les microblessures causées par les pontes de la mouche de l'olive (vecteur occasionnel). La température qui favorise l'infection va de 16 à 25 °C, avec un optimum de 22-24. Le chlorose initiée au printemps peut devenir uniforme pendant l'été. La maladie se manifeste à l'automne lors de l'invasion des fruits. L'infection est favorisée par l'humidité, spécialement surprenante et abondante dans la période végétative, particulièrement au printemps.
Les feuilles présentent des taches blanchâtres et virent au brun en se desséchant. Sur les olives, la maladie se manifeste par des taches brun-noir (spécialement au point d'insertion du pédoncule avec une dépression légèrement enfoncée de couleur brun obscur qui s'étend progressivement à tout le fruit qui se momifie et peut tomber à terre. La plante subit une défoliation plus ou moins marquée et les olives touchées ne sont plus acceptables pour la trituration. L'huile ainsi produite est un produit dégradé, rosâtre et avec une acidité (10-15%). Les olives de table sont impropres à toute confiserie.
En Italie la pathologie causée aussi par Colletotrichum acutatum concerne les régions les plus méridionales.
En septembre 2014, une attaque sans précédent sur les olives, avec également Camarosporium dalmaticum a provoqué une catastrophe économique sans précédent, attribuée à tort à la cécidiomye.
Deux modes de lutte sont possibles et complémentaires.
La prévention consiste à tailler et enlever les branches atteintes qui seront incinérées. Les olives momifiées seront récoltées et détruites dans des seaux contenant de la soude ou incinérées[3].
La lutte se fait par l'emploi de sels de cuivre sous forme de préparations diverses autorisées (bouillie bordelaise, ...)[4].
Colletotrichum gloeosporioides est une espèce de champignons ascomycètes (syn. Gleosporium olivarum). Il est l'agent de la maladie de l'anthracnose qui touche en particulier les tomates et les olives. Il appartient au genre Colletotrichum dans la famille des Glomerellaceae. Le stade sexué (téléomorphe) correspondant est Glomerella cingulata. Les espèces du genre Colletotrichum sont parmi les principaux agents pathogènes des plantes, dans le monde entier.
Colletotrichum gloeosporioides (Penz.&Sacc., 1884) (sin. Gloeosporium olivarum) è un fungo imperfetto il cui stadio perfetto è costituito dall' ascomicete Glomerella cingulata. I funghi del genere Colletotrichum, insieme con il suo stadio perfetto Glomerella, sono tra i principali agenti patogeni delle piante in tutto il mondo.
Il Colletotrichum è un genere caratterizzato dalla formazione di strutture chiamate acervuli, piatte, a forma di disco, subepidermiche, con frecce o spine; i conidiofori che lo compongono si affiancano in modo fitto sono semplici e di forma allungata, conidi unicellulari ialini, di solito a forma di bastoncino, che restano negli acervuli tenuti insieme da una massa mucillaginosa di polisaccaridi, solubile in acqua. Nonostante queste spore non siano strutture di resistenza, il micelio del fungo può rimanere vitale per lunghi periodi di tempo in semi, residui colturali, o infezioni latenti nei frutti.
Il fungo cresce rapidamente in colture quali Agar-Destrosio Patate (PDA) e forma colonie concentriche di colore bruno oliva, che possono formare settori o meno.
In quanto scatenante dell'antracnosi colpisce una gran varietà di frutta tropicale: soprattutto mango, papaya, caffè, guava, ma altresì avocado, passiflora, annona, cacao, lime; e ad altre latitudini pure fragola, melo, pesco e tra gli ortaggi il genere capsicum ed il tamarillo. Ciò naturalmente talvolta comporta danneggiamenti a carico delle rese agricole con susseguenti perdite economiche.
Il fungo, è agente della Lebbra dell'olivo. Penetra nell'organismo tramite aperture naturali (lenticelle, stomi) o microferite dovute a forature d'insetti tra cui la Mosca olearia (talvolta vettore); la temperatura che favorisce l'infezione va dai 16 ai 25 °C, con un "optimum" di 22-24. La clorosi inizia in primavera per divenire uniforme durante l'estate. La malattia si evidenzia in autunno all'invaiatura. L'infezione è favorita dall'umidità, specialmente se giunge inattesa ed abbondante nel periodo vegetativo, soprattutto in primavera.
Le foglie presentano chiazze biancastre e seccandosi virano al bruno; sulle olive, la malattia si manifesta con macchie bruno nere (spesso nel punto d'inserzione del peduncolo) e una tacca leggermente infossata di colore bruno scuro, che successivamente si estende a tutto il frutto, che mummifica e a volte cade a terra. La pianta subisce delle defoliazioni più o meno marcate e le olive colpite dalla malattia non sono più commerciabili: quelle da olio danno un prodotto scadente, rosato ad altissima acidità (10-15%), e quelle da tavola non possono essere più lavorate.
In Italia la patologia, causata anche dal Colletotrichum acutatum, interessa prevalentemente le regioni più meridionali.
Può essere Chimica: da attuare nel periodo dell'invaiatura e ripetere all'incirca dopo un mese, si effettua con trattamenti preventivi, a base di sali di rame oppure con prodotti capaci di contrastare le infezioni fungine. Biologica: Dopo un monitoraggio per decidere se intervenire, si effettua una potatura (con attrezzi ben disinfettati) per arieggiare la chioma, tagliare i rami infetti e distruggerli (così come le drupe infette) per contrastare così la diffusione della malattia; inoltre va agevolato il drenaggio dell'acqua per evitarne ristagni. Si può attuare anche una lotta integrata che contempli entrambi i metodi. Infine è preferibile non attardarsi troppo per la raccolta.
In generale i trattamenti effettuati contro l'occhio di pavone dell'olivo servono a controllare anche la lebbra delle olive.
Colletotrichum gloeosporioides (Penz.&Sacc., 1884) (sin. Gloeosporium olivarum) è un fungo imperfetto il cui stadio perfetto è costituito dall' ascomicete Glomerella cingulata. I funghi del genere Colletotrichum, insieme con il suo stadio perfetto Glomerella, sono tra i principali agenti patogeni delle piante in tutto il mondo.
Colletotrichum gloeosporioides (Penz.) Penz. & Sacc. – gatunek grzybów z rodziny Glomerellaceae[1].
Pozycja w klasyfikacji według Index Fungorum: Glomerellaceae, Incertae sedis, Hypocreomycetidae, Sordariomycetes, Pezizomycotina, Ascomycota, Fungi[1].
Po raz pierwszy takson ten zdiagnozował w 1882 r. Penzig nadając mu nazwę Vermicularia gloeosporioides. W 1884 r. ten sam autor oraz J. P.A. Saccardo nadali mu obecną, uznaną przez Index Fungorum nazwę[1].
Przez długi czas anamorfę tego gatunku traktowano jako odrębny gatunek – Glomerella cingulata (Stoneman) Spauld. & H. Schrenk 1903. Obecnie, gdy wiadomo już, że jest to tylko bezpłciowe stadium Colletotrichum gloeosporioides, nazwa Glomerella cingulata jest już tylko synonimem.
Synonimów nazwy naukowej ma ponad 80[2]:
Postać rozmnażająca się płciowo (teleomorfa) występuje rzadko. Tworzy kuliste perytecja, w których powstają jednokomórkowe i bezbarwne askospory o rozmiarach 17–22 × 3–5 μm[3].
Anamorfa w acerwulusach tworzy owalne, jednokomórkowe konidia o rozmiarach 10–115 × 5–7 μm. Pojawiają się bardzo licznie w czasie wilgotnej pogody na porażonych częściach roślin w postaci łososioworóżowych skupisk[3].
Jest szeroko rozprzestrzeniony. Występuje w różnych strefach klimatycznych na całym świecie[3].
Teleomorfę obserwowano w Polsce na pomidorze zwyczajnym, różnych gatunkach magnolii, różaneczniku katawbijskim, fiołku leśnym i fiołku skalnym[4], a także na wielu gatunkach ziół, takich, jak dzięgiel litwor, macierzanka tymianek, kminek zwyczajny, łubin Hartwega, bez czarny[5]. Anamorfę obserwowano w Polsce na jabłoniach, daktylowcu właściwym, tęgoszu mocnym, niektórych gatunkach wierzb[4], oraz na wiśni pospolitej[6].
Pasożyt występujący na wielu gatunkach roślin[5]. Wywołuje m.in. takie grzybowe choroby roślin, jak gorzka zgnilizna wiśni i antraknoza łubinu, jest też jednym z patogenów wywołujących gorzką zgniliznę jabłek[6].
Colletotrichum gloeosporioides (Penz.) Penz. & Sacc. – gatunek grzybów z rodziny Glomerellaceae.
Colletotrichum gloeosporioides é uma espécie de fungo, pertencente à ordem Melanconiales da classe Coelomycetes, cuja fase perfeita é classificado com estirpes homotálicas ou heterotálicas de ascomicetos do gênero Glomerella sp.. Os fungos deste gênero, juntamente com sua fase perfeita, são considerados os maiores patógenos de plantas em todo o mundo. [1]
O gênero Colletotrichum caracteriza-se pela formação de estruturas denominadas acérvulos, em forma de disco achatado, subepidérmico, com espinhos ou setas, conidióforos simples e alongados, conídios hialinos unicelulares, geralmente em forma de bastonete, que permanecem nos acérvulos aderidos por uma massa mucilaginosa de polissacarídeos, solúveis em água. Apesar destes esporos não serem estruturas de resistência, os micélios do fungo podem permanecer viáveis por longo período de tempo, em sementes, restos culturais, ou em infecções latentes em frutos. [2]
Tem crescimento rápido em meio de cultura Batata-Dextrose-Ágar (BDA), formando colônias concêntricas, de coloração verde-oliva à marrom, podendo ocorrer a formação ou não de setores.[carece de fontes?]
Colletotrichum gloeosporioides é uma espécie de fungo, pertencente à ordem Melanconiales da classe Coelomycetes, cuja fase perfeita é classificado com estirpes homotálicas ou heterotálicas de ascomicetos do gênero Glomerella sp.. Os fungos deste gênero, juntamente com sua fase perfeita, são considerados os maiores patógenos de plantas em todo o mundo.
Glomerella cingulata je grzib[32], co go nojprzōd ôpisoł Stoneman, a terŏźnõ nazwã doł mu Spauld. & H. Schrenk 1903. Glomerella cingulata nŏleży do zorty Glomerella i familije Glomerellaceae.[69][70][71]
Glomerella cingulata je grzib, co go nojprzōd ôpisoł Stoneman, a terŏźnõ nazwã doł mu Spauld. & H. Schrenk 1903. Glomerella cingulata nŏleży do zorty Glomerella i familije Glomerellaceae.

.JPG)



