Comprehensive Description
(
Inglês
)
fornecido por Smithsonian Contributions to Zoology
Prochilodus reticulatus Valenciennes, 1850
Prochilodus reticulatus Valenciennes in Cuvier and Valenciennes, 1850:92 [type locality: lagune de Maracaïbo (= Venezuela, Lago Maracaibo); common name],—Günther, 1864:295 [in part, Lake of Maracaibo (=Lago Maracaibo); not Kner, 1859, reference or citation of species from Brazil],—Lütken, 1875c:228 [Maracaibo-Söen (=Lago Maracaibo)].—Eigenmann, 1910: 424 [in part, not citation of species from localities outside of Lago Maracaibo basin].—Bertin, 1947:36 [syntype depository].—Fowler, 1950:222 [literature compilation, in part; not citations of occurrence of species outside of Lago Maracaibo or reported distribution outside that basin].—Mago-Leccia, 1967:250 [reported type locality of Caraccas (=Caracas, Distrito Federal, Venezuela) for Prochilodus asper Lütken, 1875, questioned and that species placed into synonymy of Prochilodus reticulatus]; 1970:31 [Venezuela]; 1972:53, fig. 7 [redescription; Venezuela, Lago Maracaibo basin; Prochilodus asper as a synonym]; 1978:5 [Venezuela].—Ringuelet et al., 1967:204 [in part, not cited occurrences of species in localities outside Lago Maracaibo basin].—Espinosa and Gimenez, 1974:1 [Venezuela, Lago Maracaibo; life history and exploitation in fisheries].—Géry, 1977:219 [Venezuela].—Taphorn and Lilyestrom, 1984a:15 [Venezuela].—Petrere, 1989:6 [efficiency of fishery for species].—Menezes and Vazzoler, 1992:62 [reproductive characteristics].—Leon et al., 1993:1 [development].—Ferrer-Montano, 1996:89 [growth, mortality, and recruitment].—Taphorn et al., 1997:79 [Venezuela].—Mojica-C, 1999:554 [Colombia, Río Catatumbo].—Institute Alexander von Humboldt, 2000:184, 189 [Colombia, Río Catatumbo basin, endangered; transplantation in Colombia].—[Not Kner, 1859:146; Perugia, 1897:25; Magalháes, 1931:111; Pearson, 1937b:109; Fowler, 1940b:98; 1950:222 [in part; not citation of species from Amazonas]; Planquette et al., 1996:132.]
Prochilodus asper Lütken, 1875c:226 (page 37 of separate) [type locality: Caraccas (= Venezuela, Distrito Federal, Caracas), cited type locality questioned by Mago-Leccia (1967:250; 1972:54), restricted herein to Venezuela, Lago Maracaibo].—Steindachner, 1879a:51 [page 35 of reprint; not cited redescription or reported presence of species in Río Magdalena].—Eigenmann and Eigenmann, 1891:48 [in listing of South American fishes].—Eigenmann, 1907b:768 [lateral-line scale count]; 1910:424 [in listing of South American fishes],—Fernández-Yépez and Martin, 1953:33 [cited type locality questioned],—Nielsen, 1974:48 [syntype depository].
Prochilodus reticulatus reticulatus.—Schultz, 1944:261 [recognition as subspecies; Venezuela, Lago Maracaibo basin; common name],—Fowler, 1975: 360 [literature compilation],—Galvis et al., 1997:37, unnumbered figures [Colombia, Lago Maracaibo basin, Río Catatumbo; common name].
Prochilodus reticulatus asper.—Schultz, 1944:261 [shift of Prochilodus asper Lütken, 1875c, to subspecies of P. reticulatus].
DIAGNOSIS.—The hyaline caudal-fin lobes of Prochilodus reticulatus discriminate that species from P. brevis, P. lacustris, P. mariae, P. nigricans, and P. rubrotaeniatus, which have dark, irregular, wavy, bar-like caudal-fin pigmentation. Among Prochilodus species with hyaline caudal fins, P. reticulatus differs from P. vimboides in the number of scales along the lateral line (41 to 45 versus 34 to 39, respectively) and in the number of horizontal rows of scales around the caudal peduncle (16 to 18 versus 13 to 15, respectively); from P. britskii in the number of horizontal rows of scales around the caudal peduncle (16 to 18 versus 13 or 14, respectively) and in the number of teeth in the inner tooth series on each side of the upper jaw (15 to 23 versus 10 to 13, respectively); from P. argenteus in the number of horizontal rows of scales between the dorsal-fin origin and the lateral line (7 to 9 versus 10 or 11), the number of vertebrae (38 to 40 versus 42 to 44, respectively), the number of scales along the lateral line (41 to 45, 43 most frequent, versus 45 to 51, 48 most frequent, respectively), and the number of median predorsal scales (11 to 18, 13 most frequent, versus 17 to 22, 18 most frequent, respectively); from P. costatus in the lack of a series of distinct, dark, wavy, horizontal stripes along the lateral surface of the body (versus the presence of such pigmentation in P. costatus) and in the number of vertebrae (38 to 40, 39 most frequent, versus 41 to 43, 42 most frequent, respectively); from P. lineatus in the lack of a series of distinct, wavy horizontal stripes along the lateral surface of the body (versus the presence of such pigmentation in P. lineatus), the number of vertebrae (38 to 40, 39 most frequent, versus 42 to 44, 43 most frequent, respectively), and the number of scales along the lateral line (41 to 45, 43 most frequent and 45 present in only 1 of 30 specimens examined for this feature, versus 44 to 50, 47 most frequent and 45 present in only 3 of 122 specimens examined for this feature, respectively); from P. hartii in the number of teeth in the inner row of the lower jaw (8 to 12 versus 13 to 18, respectively), the number of horizontal rows of scales around the caudal peduncle (16 to 18, 17 most frequent and 16 infrequent, versus 14 to 16, 14 and 15 most frequent and 16 infrequent, respectively), and the absence of wavy stripes on the sides of the body (versus the presence of such striping in all but the largest examined specimens); and from P. magdalenae in the ranges and modal values of the number of scales along the lateral line (41 to 45, 42 and 43 most frequent, 45 present in only 1 of 30 specimens examined for this feature, versus 43 to 46, 44 and 45 most frequent, respectively; Figure 48), the number of vertebrae (38 to 40, 39 most frequent, versus 39 to 42, 41 most frequent and 39 in only 8% of specimens examined for this feature, respectively; Figure 49), and the number of median predorsal scales (11 to 18, 15 and 16 most frequent, versus 12 to 17, 13 most frequent, respectively; Figure 50).
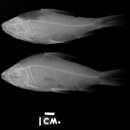
DESCRIPTION.—Morphometric and meristic data for Prochilodus reticulatus presented in Table 15. Body moderately high, transversely compressed. Greatest body depth at dorsal-fin origin. Dorsal profile of head gently concave. Predorsal profile of body moderately convex. Body profile posteroventrally inclined along dorsal-fin base; profile ranging from gently convex to straight between posterior of dorsal-fin base and adipose-fin origin, and concave along caudal peduncle. Predorsal portion of body with slight median ridge. Postdorsal portion of body obtusely rounded transversely. Ventral profile of body moderately convex from tip of lower jaw to posterior of anal-fin base. Ventral profile of caudal peduncle concave. Prepelvic region transversely flattened proximate to pelvic-fin insertion. Distinct median keel present between pelvic-fin insertion and anus.
Head profile pointed. Mouth terminal. Snout length greater than horizontal width of orbit. Nares of each side of head close to each other; anterior nares circular, posterior nares crescent shaped. Adipose eyelid present but poorly developed; most developed anteriorly but with greater part of eye uncovered. Lips fleshy, moderately developed, and forming oral disk when protracted.
Functional teeth in two rows in each jaw. All teeth movably implanted in flesh that overlies jaws. All teeth of similar size, with exposed portions spoon shaped except when worn down. Inner tooth series in each jaw with 15 to 23 teeth on left side of upper jaw and 8 to 12 teeth on left side of lower jaw. Outer row of teeth in each jaw with approximately 103 teeth on each side of upper jaw and approximately 80 teeth on each side of lower jaw in lectotype of P. asper (tooth counts in dried lectotype and paralectotypes of P. reticulatus impossible to determine). Upper and lower lips bordered by numerous globular, fleshy papillae.
Scales spinoid. Scales in middorsal series between posterior of dorsal-fin base and adipose-fin origin similar in form to those of adjoining portions of body. Lateral line with 41 to 45 (70% of specimens with 43 or 44) scales; 7 to 9 (80.0% of specimens with 8) horizontal rows of scales between dorsal-fin origin and lateral line; 7 or 8 (73.3% of specimens with 7) horizontal rows of scales between pelvic-fin insertion and lateral line; 6 or 7 (60.0% of specimens with 7) horizontal rows of scales between anal-fin origin and lateral line; 11 to 18 (34.0% of specimens with 13) median predorsal scales; 12 to 16 (48.3% of specimens with 14) scales in middorsal series between posterior of dorsal-fin base and adipose-fin origin; 16 to 19 (56.7% of specimens with 18) horizontal rows of scales around caudal peduncle.
Dorsal fin preceded by small, but well-developed, anteroventrally bifurcate, procumbent spine somewhat triangular in lateral view. Dorsal-fin rays (including procumbent spine) iii,10 [iii,10]; anal-fin rays iii,7 to 9 (iii,8 most frequent) [iii,8]; pectoral-fin rays i,13 to 15 (i,14 most frequent) [i,14]; pelvic-fin rays i,8 [i,8]; principal caudal-fin rays 10/9 [10/9].
Vertebrae 38 to 40 (59.5% of specimens with 39).
Dorsal fin truncate, slightly pointed distally; posterior unbranched and anterior branched rays longest and subequal. Dorsal-fin origin located closer to tip of snout than to caudal- fin base. Greatest length of adipose fin approximately equal to horizontal width of orbit. Adipose-fin origin located along vertical that passes through middle of anal-fin base. Pectoral fin distally pointed. Tip of adpressed pectoral fin reaching posteriorly almost to pelvic-fin insertion. Pelvic fin falcate. Pelvic-fin insertion typically located along vertical that passes though anterior one-fourth of dorsal-fin base. Tip of adpressed pelvic fin reaching, or almost reaching, anus. Axillary scale present, its length approximately one-fourth of greatest length of pelvic fin. Posterior unbranched and anterior branched anal-fin rays longest and subequal. Caudal fin bifurcate.
COLORATION IN ALCOHOL.—Ground coloration silvery yellow, with dorsal portions of body and head darker. Lateral surface of body with up to 12 dark, vertically elongate, diffuse and irregular patches of pigmentation between head and caudal fin. Patches with approximate overall form of narrow isosceles triangles with apexes located along middle of ventrolateral portion of body and bases along dorsomedial region of body. Pigment patches well developed in small specimens, but indistinct or absent in large individuals. Pattern of distinct, wavy, dark, horizontal stripes present on lateral surface of body in many congeners not apparent in examined specimens. Field of black or brown chromatophores forming dark, irregular spot on upper one-half of opercle.
Dorsal fin with 3 to 8 (most frequently 6) dark, irregular stripes beginning on anterior margin of fin and running approximately parallel to dorsal-fin base. Adipose fin with dorsal margin finely bordered with black. Pectoral, pelvic, anal, and caudal fins dusky. Iris yellowish silver, with diffuse dusky dorsal and ventral regions.
COLORATION IN LIFE.—According to Schultz (1944:264), the dorsal fin is pinkish and the sides of the body are yellowish.
DISTRIBUTION.—Prochilodus reticulatus is endemic to the rivers of the Lago Maracaibo basin in northwestern Venezuela and northeastern Colombia (Figure 52, triangles).
COMMON NAME.—Bocachico de Maracaibo (Venezuela).
BIOLOGY AND FISHERIES.—Espinosa and Gimenez (1974) discussed the biology of this species and reported that in the southwestern portion of the Lago Maracaibo basin Prochilodus reticulatus is heavily exploited in commercial fisheries.
COMPARISONS.—Differences in various pigmentation and meristic features unequivocally distinguish Prochilodus reticulatus of the Lago Maracaibo basin from all of its congeners except for P. magdalenae, the only other Prochilodus species with a Trans-Andean distribution. The two nominal forms differ in the ranges and modal values of the number of scales along the lateral line, the number of medial predorsal scales, and the number of vertebrae (see “Diagnosis,” above, and Figures 48 to 50). In light of these differences and their disjunct areas of occurrence, these two nominal forms are treated as separate species herein.
MATERIAL EXAMINED.—77 specimens (30, 64.6–278.6 mm SL; partial meristic data taken from additional 25 specimens).
VENEZUELA. Trujillo: Rio Jimelles, tributary Rio Motatan, 12 km E of Motatán, USNM 121332, 1 (1, 201.5) [1R]. Rio Motatán, bridge 22 km N of Motatán, USNM 121327, 1 (1, 175.1) [1R]. Zulia: Encontrados, FMNH 6335, 3 (1, 171.3191.4) [1R]. Lago Tulé, approximately 75 km W of Maracaibo, 5 km from Rio Socuy, USNM 121330, 2 (2, 130.5–157.5) [2R]. Rio Socuy, approximately 3 km upriver of its mouth, USNM 121328, 9 (4, 64.6–203.5) [2R]. Small lagoon along Rio San Ignacio, approximately 20 km S of Rosario, USNM 121326, 4 (2, 110.9–115.2) [2R], Rio Apón, approximately 35 km S of Rosario, USNM 121331, 6 (4, 198.3–246.5) [4R]. Caño 0.5 mile (0.8 km) W of Sinamaica, USNM 121329, 2 (2, 203.8212.8) [2R]. Rio Palmar near Totumo, approximately 100 km SW of Maracaibo, USNM 121324, 31 (2, 118.2–243.5) [15R]. Rio Negro, near to its confluence with Rio Yasa, USNM 121333, 10 (3, 106.0–219.0) [10R]. Inexact Locality: Lago Maracaibo, MNHN A.8637–38, 2 (2, 244.3–248.2, paralectotypes of Prochilodus reticulatus); MNHN A.8639, 1 (1, 278.6, lectotype of Prochilodus reticulatus); MNHN A.8640, 1 (264.8, paralectotype of Prochilodus reticulatus). “Caracas” (probably from Lago Maracaibo, see “Remarks,” above), BMNH 1876.1.10:25, 1 (165.1); NMW 56635, 1 (1, 212.4; possible paralectotype of Prochilodus reticulatus, see “Remarks,” above) [1R]; ZMUC 72, 1 (1, 241.5, paralectotype of Prochilodus asper) [1R]; ZMUC 73, 1 (191.5, lectotype of Prochilodus asper) [1R].
- citação bibliográfica
- Castro, Ricardo M. C. and Vari, Richard P. 2004. "Detritivores of the South American fish family Prochilodontidae (Teleostei:Ostariophysi:Characiformes) : a phylogenetic and revisionary study." Smithsonian Contributions to Zoology. 1-189. https://doi.org/10.5479/si.00810282.622