Das FSME-Virus (FSMEV, wissenschaftlich Tick-borne encephalitis virus, TBEV) ist eine Spezies behüllter einzelsträngiger RNA-Viren positiver Polarität und der Erreger der Frühsommer-Meningoenzephalitis (FSME bzw. TBE). Es handelt sich um ein humanpathogenes Virus aus der Familie der Flaviviridae, Gattung Flavivirus.
Es werden drei Subtypen unterschieden[3].
Die Typuslinie ist der Sofjin strain.[5]
Das Viruskapsid besteht aus drei Strukturproteinen:
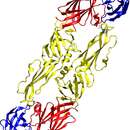
Das Glykoprotein E spielt eine zentrale Rolle in der Biologie der Infektion und ist für die Bindung und das Eindringen in die Zielzelle verantwortlich. Es gehört zu den am besten charakterisierten viralen Proteinen überhaupt.
Zecken (Ixodida sp.) sind die Hauptvektoren und das Hauptreservoir der FSME. Die Ansteckung erfolgt über Zeckenstiche oder über Rohmilch infizierter Tiere. Die Krankheit selbst wurde erstmals 1931 bei Forstarbeitern aus Neunkirchen beschrieben, der Erreger 1949 isoliert.
Das endemische Auftreten von FSME ist immer mit großen Flüssen assoziiert. Die Gründe dafür sind bis dato unklar.
Die Spezies TBEV gehört innerhalb der Gattung Flavivirus zum so genannten Tick-Borne-Enzephalitis-Komplex (TBE)[6], zu dem die neben diesen Erregern der Frühsommer-Meningoenzephalitis auch das Louping-Ill-Virus (Erreger von Louping III, LI), das Kyasanur-Forest-Disease-Virus (Erreger des Kyasanur-Wald-Fiebers, KFD) mit dem Subtyp Al-Khurma-Virus, das Powassan-Virus (Erreger der Powassan-Virus-Enzephalitis, PE), das Omsk-hämorrhagisches-Fieber-Virus (Erreger des Omsker Fiebers), das Langat-Virus, sowie die Erreger der Negeshivirus-Enzephalitis zählen. Die einzelnen Arten sind jeweils auf bestimmte Regionen begrenzt.
In Deutschland ist der direkte oder indirekte Nachweis des FSME-Virus namentlich meldepflichtig nach des Infektionsschutzgesetzes (IfSG), soweit der Nachweis auf eine akute Infektion hinweist. Die Meldepflicht betrifft in erster Linie die Leitungen von Laboren ( IfSG).
In der Schweiz ist der positive und negative laboranalytische Befund zu einem Zeckenenzephalitisvirus für Laboratorien meldepflichtig und zwar nach dem Epidemiengesetz (EpG) in Verbindung mit der Epidemienverordnung und der Verordnung des EDI über die Meldung von Beobachtungen übertragbarer Krankheiten des Menschen.
Das FSME-Virus (FSMEV, wissenschaftlich Tick-borne encephalitis virus, TBEV) ist eine Spezies behüllter einzelsträngiger RNA-Viren positiver Polarität und der Erreger der Frühsommer-Meningoenzephalitis (FSME bzw. TBE). Es handelt sich um ein humanpathogenes Virus aus der Familie der Flaviviridae, Gattung Flavivirus.

FSME wiiren
Abb. A: pH=8,0
Abb. B: pH=10,0
Abb. C: pH=5,4
At FSME-wiirus suragt för meningoenzephalitis an hiart tu't famile Flaviviridae.
Hat woort faan tegen (Ixodida slacher) auerdraanj.
At FSME-wiirus suragt för meningoenzephalitis an hiart tu't famile Flaviviridae.
Tick-borne encephalitis virus (TBEV) is a positive-strand RNA virus associated with tick-borne encephalitis in the genus Flavivirus.
TBEV is a member of the genus Flavivirus. Other close relatives, members of the TBEV serocomplex, include Omsk hemorrhagic fever virus, Kyasanur Forest disease virus, Alkhurma virus, Louping ill virus and Langat virus.[1]
TBEV has three subtypes:
The reference strain is the Sofjin strain.[3]
TBEV is a positive-sense single-stranded RNA virus, contained in a 40-60 nm spherical, enveloped capsid.[4] The TBEV genome is approximately 11kb in size, which contains a 5' cap, a single open reading frame with 3' and 5' UTRs, and is without polyadenylation.[4] Like other flaviviruses,[5] the TBEV genome codes for ten viral proteins, three structural, and seven nonstructural (NS).[6][4]
The structural proteins are C (capsid), PrM (premembrane, which is cleaved to produce the final membrane protein, M), and E (envelope). The seven nonstructural proteins are: NS1, NS2A, NS2B, NS3, NS4A, NS4B, and NS5. The role of some nonstructural proteins is known, NS5 serves as RNA-dependent RNA polymerase, NS3 has protease (in complex with NS2B) and helicase activity.[6][4] Structural and nonstructural proteins are not required for the genome to be infectious.[4] All viral proteins are expressed as a single large polyprotein, with the order C, PrM, E, NS1, NS2A, NS2B, NS3, NS4A, NS4B, NS5.[6]
Infection of the vector begins when a tick takes a blood meal from an infected host. This can occur at any part of the tick's life cycle but a horizontal transmission between infected nymphs and uninfected larvae co-feeding on the same host is thought to be key in maintaining the circulation of TBEV.[7][4] TBEV in the blood of the host infects the tick through the midgut, from where it can pass to the salivary glands to be passed to the next host. In non-adult ticks, TBEV is transmitted transtadially by infecting cells that are not destroyed during molting, thus the tick remains infectious throughout its life.[7] Infected adult ticks may be able to lay eggs that are infected, transmitting the virus transorvarially.[8]
In humans, the infection begins in the skin (with the exception of food-borne cases, about 1% of infections) at the site of the bite of an infected tick, where Langerhans cells and macrophages in the skin are preferentially targeted.[6] TBEV envelope (E) proteins recognize heparan sulfate (and likely other receptors) on the host cell surface and are endocytosed via the clathrin mediated pathway. Acidification of the late endosome triggers a conformational change in the E proteins, resulting in fusion, followed by uncoating, and release of the single-stranded RNA genome into the cytoplasm.[9][4]
The viral polyprotein is translated and inserts into the ER membrane, where it is processed on the cytosolic side by host peptidases and in the lumen by viral enzyme action. The viral proteins C, NS3, and NS5 are cleaved into the cytosol (though NS3 can complex with NS2B or NS4A to perform proteolytic or helicase activity), while the remaining nonstructural proteins alter the structure of the ER membrane. This altered membrane permits the assembly of replication complexes, where the viral genome is replicated by the viral RNA-dependent RNA polymerase, NS5.[9][6]
Newly replicated viral RNA genomes are then packaged by the C proteins while on the cytosolic side of the ER membrane, forming the immature nucleocapsid, and gain E and PrM proteins, arranged as a heterodimer, during budding into the lumen of the ER. The immature virion is spiky and geometric in comparison to the mature particle. The particle passes through the golgi apparatus and trans-golgi network, under increasingly acidic conditions, by which the virion matures with cleavage of the Pr segment from the M protein and formation fusion competent E protein homodimers. Though the cleaved Pr segment remains associated with protein complex until exit.[4][9]
The virus is released from the host cell upon fusion of the transport vesicle with the host cell membrane, the cleaved Pr now segments dissociate, resulting in a fully mature, infectious virus.[4][9] However, partially mature and immature viruses are sometimes released as well; immature viruses are noninfectious as the E proteins are not fusion competent, partially mature viruses are still capable of infection.[9]
With the exception of food-borne cases, infection begins in the skin at the site of the tick bite. Skin dendritic (or Langerhans) cells (DCs) are preferentially targeted.[6] Initially, the virus replicates locally and immune response is triggered when viral components are recognized by cytosolic pattern recognition receptors (PRRs), such as Toll-like receptors (TLRs).[10] Recognition causes the release of cytokines including interferons (IFN) α, β , and γ and chemokines, attracting migratory immune cells to the site of the bite.[6] The infection may be halted at this stage and cleared, before the onset of noticeable symptoms. Notably, tick saliva enhances infection by modulating host immune response, dampening apoptotic signals.[10] If the infection continues, migratory DCs and macrophages become infected and travel to the local draining lymph node where activation of polymorphonuclear leukocytes, monocytes and the complement system are activated.[10]
The draining lymph node can also serve as a viral amplification site, from where TBEV gains systemic access. This viremic stage corresponds to the first symptomatic phase in the prototypical biphasic pattern of tick-borne encephalitis.[4] TBEV has a strong preference for neuronal tissue, and is neuroinvasive.[11] The initial viremic stage allows access to a number of the preferential tissues. However, the exact mechanism by which TBEV crosses into the central nervous system (CNS) is unclear.[11][10][8][4] There are several proposed mechanism for TBEV breaching the blood-brain barrier (BBB): 1)The "Trojan Horse" mechanism, whereby TBEV gains access to the CNS while infecting an immune cell that passes through the BBB;[10][6][11] 2) Disruption and increased permeability of the BBB by immune immune cytokines;[11] 3) Via infection of the olfactory neurons;[6] 4) Via retrograde transport along peripheral nerves to the CNS;[6] 5) Infection of the cells that make up part of the BBB.[6][10]
CNS infection brings on the second phase in the classic biphasic infection pattern associated with the European subtype. CNS disease is immunopathological; release of inflammatory cytokines coupled with the action of cytotoxic CD8+ T cells and possibly NK cells results in inflammation and apoptosis of infected cells that is responsible for many of the CNS symptoms.[10][11]
TBEV specific IgM and IgG antibodies are produced in response to infection.[4] IgM antibodies appear and peak first, as well as reaching higher levels, and typically dissipate in about 1.5 months post infection, though there exists considerable variation from patient to patient. IgG levels peak at about 6 weeks after the appearance of CNS symptoms, then decline slightly but do not dissipate, likely conferring life long immunity to the patient.[4][6]
The ancestor of the extant strains appears to have separated into several clades approximately 2750 years ago.[12] The Siberian and Far Eastern subtypes diverged about 2250 years ago. A second analysis suggests an earlier date of evolution (3300 years ago) with a rapid increase in the number of strains starting around 300 years ago.[13] Different strains of the virus have been transmitted at least three times into Japan between 260–430 years ago.[14][15] The strains circulating in Latvia appear to have originated from both Russia and Western Europe[16] while those in Estonia appear to have originated in Russia.[17] The Lithuanian strains appear to be related to those from Western Europe.[18] Phylogenetic analysis indicates that the European and Siberian TBEV sub-types are closely related while the Far-eastern sub-type is closer to the Louping Ill Virus.[1] However, in antigenic relatedness, based on the E, NS3, and NS5 proteins, all three sub-types are highly similar, and Louping Ill virus is the closest relative outside the collective TBEV group.[19]
Though the first description of what may have been TBE appears in records in the 1700s in Scandinavia,[11] identification of the TBEV virus occurred in the Soviet Union in the 1930s.[20] The investigation began due to an outbreak of what was believed to be Japanese Encephalitis ("Summer encephalitis"), among Soviet troops stationed along the border with the Japanese empire (present day People's Republic of China), near the Far Eastern city of Khabarovsk. The expedition was led by virologist Lev A. Zilber, who assembled a team of twenty young scientists in a number of related fields such as acarology, microbiology, neurology, and epidemiology.[21][20] The expedition arrived in Khabarovsk on May 15, 1937, and divided into squads, Northern-led by Elizabeth N. Levkovich and working in the Khabarovski Krai- and Southern-led by Alexandra D. Sheboldaeva, working in the Primorski Krai.[20]
Inside the month of May, the expedition had identified ticks as the likely vector, collected I. persucatus ticks by exposure of bare skin by entomologist Alexander V. Gutsevich and virologist Mikhail P. Chumakov had isolated the virus from ticks feeding on intentionally infected mice. During the summer, five expeditions members became infected with TBEV, and while there were no fatalities, three of the five suffered damaging sequelae.[20]
The expedition returned in mid-August and in October of 1937 Zilber and Sheboldova were arrested, falsely accused of spreading Japanese encephalitis. Expedition epidemiologist Tamara M. Safonov, was arrested the following January for protesting the charges against Zilber and Sheboldova. As a consequence of the arrests, one of the important initial works was published under the authorship of expedition acarologist, Vasily S. Mironov. Zilber was released in 1939 and managed to restore, along with Sheboldova, co-authorship on this initial work; however, Safanov and Sheboldova (who was not released) spent 18 years in labor camps.[20][21]
Tick-borne encephalitis virus (TBEV) is a positive-strand RNA virus associated with tick-borne encephalitis in the genus Flavivirus.
El virus de la encefalitis por garrapatas (TBEV, Tick-Borne Encephalitis-virus) es un virus ARN monocatenario positivo transmitido por garrapatas, del género Flavivirus, de la familia Flaviviridae, grupo IV orden sin clasificar, que causa la encefalitis transmitida por garrapatas.[1]
El virus de la encefalitis por garrapatas (TBEV, Tick-Borne Encephalitis-virus) es un virus ARN monocatenario positivo transmitido por garrapatas, del género Flavivirus, de la familia Flaviviridae, grupo IV orden sin clasificar, que causa la encefalitis transmitida por garrapatas.
Puukentsefaliidi viirus ( tick-borne encephalitis virus;lüh TBEV) on viiruseliik Flaviviridae sugukonnas.
Puukentsefaliidi viirus liigitatakse RNA-viiruste hulka ja sellel tunnistatakse järgmiseid alatüüpe:
Viirust seostatakse puukentsefaliidiga.
Siberi alam-tüüpi viirus võib põhjustada puugihammustuse järgselt vastuvõtlikel inimestel Siberi alatüübi puukentsefaliidi vormi Siberi entsefaliiti, mis võib lõppeda surmaga ja millel ei ole veel ravi leitud.
Siberi entsefaliidi juhtumeid on diagnoositud Soomes kokku ligi 20 ning on esinenud ka surmajuhtumeid.[1]
Puukentsefaliidi viirus ( tick-borne encephalitis virus;lüh TBEV) on viiruseliik Flaviviridae sugukonnas.
Le Virus de la méningoencéphalite à tiques, VMET (en anglais : Tick-borne Encephalitis Virus, TBEV) est un virus à ARN monocaténaire de polarité positive (groupe IV de la classification Baltimore) appartenant à la famille des Flaviviridae et au genre Flavivirus, principalement responsable de l'encéphalite à tiques.
Dans la même famille on trouve également le virus de la fièvre hémorragique d'Omsk, celui de la fièvre de Kyasanur, le virus Alkhurma, le virus louping ill (responsable de l’encéphalite virale ovine[2]) et le virus de Langat.
Le VMET est endémique sur le nord de l'Eurasie (de la zone frontalière de l'est de la France à l'Europe centrale, de l'Ouest de la Sibérie jusqu'en Extrême Orient et au Japon). Le nombre de cas d'encéphalites à tiques est estimé à près de dix mille cas mondiaux par an.
Il s’agit d’un virus enveloppé à ARN simple brin non-segmenté de sens positif et de symétrie icosaédrique.
Son génome code une polyprotéine de structure (SP) et sept protéines non structurales[3]. Le virus EBV est sphérique, son diamètre est de 50 nm, soit 1/20e de micromètre, il est constitué d'une enveloppe virale qui protège un virion d'ARN associé à une protéine structurale « C ». L'enveloppe externe est porteuse de deux protéines, « M » et « E », la seconde étant impliquée dans l'immunogénicité et l'identification du virus.
Son ARN est séquencé depuis 1989.
L'assemblage des virus EBV est complexe, se déroulant en plusieurs stades de maturation. La plupart de ces étapes ont été étudiées sur des cellules de mammifères, et l'on a peu de données sur celles de tiques[3].
Les virions matures s'attachent aux récepteurs de membrane de la cellule cible, et la pénètrent le plus souvent par endocytose. La réplication du génome s'effectue dans les membranes du réticulum endoplasmique, et les nouvelles protéines virales sont synthétisées par les ribosomes.
Les virions immatures sont transportés dans l'appareil de Golgi pour effectuer leur maturation (changement de structure de la protéine M). Cette maturation n'est pas toujours complète, ce qui amène à la production de virions à différents stades : immatures, partiellement matures et complètement matures. Les virions immatures ne sont pas infectieux (incapables de fusion avec une cellule cible), mais les virions matures (en partie ou complètement) peuvent entamer un nouveau cycle infectieux[3].
La production de formes partiellement matures (hétérogènes du point de vue structurel et antigénique) a été interprétée comme une stratégie d'échappement immunitaire, et une façon de s'adapter à de nouvelles cellules-cibles (de tissus plus variés)[3].
Le VMET est endémique sur le nord de l'Eurasie (zone frontalière de la région de Strasbourg à l'Europe centrale, et jusqu'au Japon en passant par la Sibérie).
Les réservoirs de virus sont les tiques elles-mêmes (transmission verticale de la femelle à sa descendance), et de nombreuses espèces de mammifères sauvages (petits rongeurs, renards, sangliers, cerfs..) et domestiques (chèvres, vaches, moutons et chiens)[4].
Le virus VMET est habituellement transmis par piqûre de tiques infectées du genre Ixodes, mais il existe aussi une contamination par produits laitiers non pasteurisés provenant d'animaux domestiques infectés[3].
En Europe, la prévalence du virus chez les tiques Ixodes ricinus varie entre 0,1 et 1,2% selon les pays et les zones étudiées[4]. Le nombre de cas humains d'encéphalites à tiques est estimé à près de dix mille cas mondiaux par an[3], ou de 5 000 à 13 000 cas[5].
L'incidence des encéphalites à tiques est en augmentation, probablement liée au réchauffement climatique (surabondance de tiques par allongement de leur période d'activité et extension de leur habitat)[3].
Il existe un vaccin efficace, mais trop peu utilisé par les groupes à risques[3]. En France, cette vaccination n'est pas recommandée pour les résidents (la maladie est très rare en France), elle peut l'être pour les voyageurs séjournant, au printemps et en été, en zone forestière d'Europe centrale et de l'est[5].
Le VMET possède trois sous-types (caractéristiques cliniques et épidémiologiques)[3],[6],[7] :
Les variations génétiques entre ces sous-types sont faibles (de 2,2% à 5,6% pour la séquence d'acides aminés de la protéine de structure), ce qui pourrait aider à déterminer des facteurs de virulence expliquant les différences cliniques[3].
Malgré les progrès réalisés au début du XXIe siècle par la recherche sur les flavivirus, et une meilleure compréhension du VEMT, il reste encore de nombreuses inconnues sur son cycle de vie. En particulier, les facteurs des virulence responsables des différences cliniques entre sous-types de virus n'ont pas été identifiés, de même, le développement du virus chez les tiques a été peu étudié[3].
Le Virus de la méningoencéphalite à tiques, VMET (en anglais : Tick-borne Encephalitis Virus, TBEV) est un virus à ARN monocaténaire de polarité positive (groupe IV de la classification Baltimore) appartenant à la famille des Flaviviridae et au genre Flavivirus, principalement responsable de l'encéphalite à tiques.
Dans la même famille on trouve également le virus de la fièvre hémorragique d'Omsk, celui de la fièvre de Kyasanur, le virus Alkhurma, le virus louping ill (responsable de l’encéphalite virale ovine) et le virus de Langat.
Le VMET est endémique sur le nord de l'Eurasie (de la zone frontalière de l'est de la France à l'Europe centrale, de l'Ouest de la Sibérie jusqu'en Extrême Orient et au Japon). Le nombre de cas d'encéphalites à tiques est estimé à près de dix mille cas mondiaux par an.
Virus krpeljnog meningoencefalitisa je RNK virus uzročnik bolesti krpeljni meningoencefalitis, koji pripada porodici flavivirusa (lat. Flaviviridae).
Serološki se mogu razlikovati dva podtipa virusa krpeljnog meningoencefalitisa: srednjoeuropski podtip i dalekoistočni podtip. Oba tipa mogu uzrokovati bolest.
![]() Nedovršeni članak Virus krpeljnog meningoencefalitisa koji govori o biologiji treba dopuniti. Dopunite ga prema pravilima Wikipedije.
Nedovršeni članak Virus krpeljnog meningoencefalitisa koji govori o biologiji treba dopuniti. Dopunite ga prema pravilima Wikipedije.
I virus della encefalite trasmessa da zecche (Tick-borne encephalitis virus, TBEV) sono una specie di arbovirus della famiglia Flaviviridae, genere Flavivirus, appartiene al IV gruppo dei virus a ((+) ssRNA). Esso è associato alla encefalite virale trasmessa dalle zecche.
Il virus fa parte dei flavivirus patogeni per l'uomo.[1]
Questo virus ha un genoma a singolo filamento di RNA positivo ((ss) (+) RNA], esso può essere distinto in tre sottotipi:[2]
Il capside virale è composto di tre proteine strutturali :
La glicoproteina E svolge un ruolo centrale nella biologia delle infezioni ed è responsabile del legame e della penetrazione nella cellula bersaglio.
Altri virus assimilabili trasmessi da zecche sono:
Le zecche vettore appartengono tipicamente a generi della famiglia Ixodidae (zecche dure):
Le zecche oltre che vettori sono anche serbatoi di riserva del virus in grado di garantirne la persistenza nell’ambiente oltre il periodo di attività della zecca tramite la trasmissione transovarica (l'infezione si trasmette attraverso le uova dalla madre alla prole), transtadiale (da larva a ninfe e / o da ninfe ad adulto), per co-feeding (zecca che effettua il pasto di sangue accanto ad un’altra infetta)i . L'uomo è il principale ospite terminale insieme a piccoli animali selvatici, per lo più roditori, che sono ospiti comuni.[3][4]
I piccoli mammiferi come i roditori sono ospiti e danno una virus amplificazione per il loro alto tasso riproduttivo. Gli esseri umani sono infettati sporadicamente, sia da una puntura di zecca o per l'ingestione di latte non pastorizzato o suoi prodotti lattiero-caseari.[5][6][7][8][9] Altri mammiferi (ad esempio ruminanti) possono anche essere infettati, ma il più delle volte non mostrano segni clinici.[10]
I virus provocano encefaliti, con una fase iniziale con febbre, mal di testa e mialgia. Nei casi più gravi segue dopo circa una settimana dalla apparente guarigione clinica una seconda fase della malattia con meningoencefalite o mielite. Questa ultima tende a causare paralisi flaccida dell'arto superiore e della spalla dei muscoli respiratori. Il coinvolgimento bulbare (tronco cerebrale) porta ad insufficienza respiratoria e quindi la morte.[3] Si ritrova nel liquido cerebrospinale la presenza di neutrofili, con una leucocitosi periferica indicativo di meningite.[3]
I virus della encefalite trasmessa da zecche (Tick-borne encephalitis virus, TBEV) sono una specie di arbovirus della famiglia Flaviviridae, genere Flavivirus, appartiene al IV gruppo dei virus a ((+) ssRNA). Esso è associato alla encefalite virale trasmessa dalle zecche.
Il virus fa parte dei flavivirus patogeni per l'uomo.
Virus klopnega meningoencefalitisa (virus KME) je flavivirus, ki ga prenašajo klopi[1] in povzroča klopni meningoencefalitis. V naravi kroži virus med klopi, ki so glavni prenašalci virusa, in gozdnimi sesalci, ki so glavni gostitelji virusa.[2] V Sloveniji so virus našli pri okoli 0,5 % klopov in 6 % malih glodavcev (v večjem deležu pri rovkah kot pri miših).[3]
Poznamo tri podtipe virusa KME:[3][4]
Virusi KME so okrogli enovijačni RNK-virusi iz rodu Flavivirus in družine Flaviviridae. Genom nosi zapis za tri strukturne beljakovine (C, prM in E) in sedem nestrukturnih beljakovin (NS1, NS2A, NS2B, NS3, NS4A, NS4B, NS5).[3][5] Beljakovina C (angl. capside protein) tvori virusno kapsido, ta pa je obdana z lipidnim dvoslojem, ki vsebuje beljakovini prM (precursor M protein) in E (envelope protein). Beljakovina E predstavlja najpomembnejši virusni antigen in sodeluje pri vezavi virusa na (še neznan) receptor na gostiteljevi celici ter vstop vanjo.[5]
Virus klopnega meningoencefalitisa (virus KME) je flavivirus, ki ga prenašajo klopi in povzroča klopni meningoencefalitis. V naravi kroži virus med klopi, ki so glavni prenašalci virusa, in gozdnimi sesalci, ki so glavni gostitelji virusa. V Sloveniji so virus našli pri okoli 0,5 % klopov in 6 % malih glodavcev (v večjem deležu pri rovkah kot pri miših).


