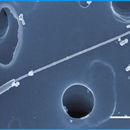
Micromonas is a genus of green algae in the family Mamiellaceae.[1][2]
Before the characterization in 2016 of a second species, Micromonas commoda, Micromonas pusilla was considered to be the only species in the genus,[3][4] which led to a disproportionate amount of research discussing a single species within the genus. It is suggested to be the dominant photosynthetic picoeukaryote in some marine ecosystems.[5] Unlike many marine algae, it is distributed widely in both warm and cold waters.[6] It is a strong swimmer and exhibits a phototaxic response.[6]
Micromonas pusilla is divided into 3 to 5 different clades despite their similarity in morphologies and habitats.[7][8] Varying ratios of clades contribute to the M. pusilla population throughout the marine ecosystem leading to the hypothesis of clades arising based on niche occupation and susceptibility to virus infection.[8]
Micromonas pusilla is considered the first picoplankton studied, when it was discovered and named Chromulina pusilla in the 1950s by R. Butcher.[9] Later, electron micrographs by the English scientists, Irene Manton and Mary Park, in the 1960s provided further details on M. pusilla.[9]
Micromonas is a group of small unicellular pear-shaped micro-algae that do not have a cell wall.[10][11][12] Just like other members in the class, they have a single mitochondrion and a single chloroplast,[12] which covers almost half of the cell.[12][13] They are able to swim due to the presence of a scale-less flagellum.[12][13][3] The axonemal structure of the flagellum for this genus is different in that the peripheral microtubules do not extend up to the termination of the central pair of microtubules, allowing a visible investigation of the motion of the central pair.[11][14][15] In Micromonas, the central pair constantly rotates in an anti-clockwise direction, despite the motion of other components of the flagellum.[11][14]
While the cell size, shape and the location of insertion of the flagellum into the cell are similar among strains and genetic clades, the variation in respective hair point length results in different lengths of the flagella within the genus.[3]
The antibiotic susceptibility was determined using a single strain of M. pusilla with the purpose to produce axenic cultures to be used in studies and experiments.[16] The strain of M.pusilla was tested with a range of antibiotics to determine the possible effects of the particular antibiotic.[16]
Resistance:[16] benzylpenicillin, gentamicin, kanamycin, neomycin, streptomycin
Sensitive:[16] chloramphenicol, polymyxin B
For M. pusilla, sensitivity towards an antibiotic is likely defined by the impairment of growth, rather than a lethal effect, when subjected to bactericidal levels of that particular antibiotic.[16] The susceptibility of other strains of M. pusilla towards this set of antibiotics should be the same.[16]
Micromonas diverged early on from the lineage that led to all modern terrestrial plants. Individual species have very similar 18S ribosomal RNA gene sequences, a comparison often used to determine microscopic speciation, however, only 90% of different genes are shared between all Micromonas species. This presence or absence of complete genes, compared with only small changes of sequence to specific genes, suggests that Micromonas is the result of intensive horizontal gene transfer.[10]
The original Micromonas reference genome was created from a strain, RCC299, first isolated in 1998 from an Equatorial Pacific sample. This strain has been continuously cultured for two decades and is available from the Roscoff Culture Collection. In 2005, a monoclonal culture of the strain was isolated. The axenic strain is available from the Center for Culture of Marine Phytoplankton, under the name CCMP2709. Currently, a separate stain that was isolated from temperate coastal waters is being sequenced.[10]
The entire Micromonas sp. genome was first shotgun sequenced in 2014. Micromonas has about 19Mb, however this varies slightly between species and strain. This is made up of 17 chromosomes and contains 59% GC content.[17] Based on [open reading frames] the genome codes for about 10000 proteins and 70 functional RNAs.[18]
Micromonas reproduces asexually through fission.[11] It has been observed that M. pusilla shows variability in optical characteristics, for example cell size and light scattering, throughout the day.[19] There is an increase in these measurements during the period with light, followed by a decrease during period without light.[19][20] This coincides with the findings that proteomic profiles change over the diel cycle, with an increase in expression of proteins related to cell proliferation, lipid and cell membrane restructuring in the dark when cells start dividing and become smaller.[20] However, the expression levels of genes and proteins can still vary within the same metabolic pathway.[20] It has also been suggested that the structure of 3’ UTR may play a role in the regulatory system.[20]
Micromonas species still share the same collection of photosynthetic pigments as the members of the class Mamiellophyceae,[3] which includes the common pigments chlorophyll a and chlorophyll b,[21] as well as prasinoxanthin (xanthophyll K), the first algal carotenoid being assigned with a structure that has a γ-end group.[22] It has been discovered that most of its xanthophylls are in the oxidized state and show similarities to ones possessed by other important marine planktons like diatoms, golden and brown algae, and dinoflagellates.[23] In addition, there is another pigment called Chl cCS-170 can be found in some strains of Micromonas and Ostreococcus living in deeper part of the ocean, which may indicate a potential adaptation for organisms that reside under low light intensity.[3]
The light-harvesting complexes of Micromonas are distinguishable from other green algae in terms of pigment composition and stability under unfavorable conditions.[21] It has been showed that these proteins use three different pigments for light harvesting, and they are resistant to high temperature and the presence of detergent.
Even though the chloroplasts, which are suggested to be originated from Cyanobacteria via endosymbiosis,[24] from Micromonas do not have a surrounding peptidoglycan layer, the peptidoglycan biosynthesis pathway is found to be complete in M. pusilla and partial in M. commoda, with the presence of some relevant enzymes only.[12] While the role of this pathway for Micromonas is still under investigation, this observation shows a lineage for different species of Micromonas along with glaucophyte algae which still have their chloroplasts covered with peptidoglycan.[12]
Micromonas make up a significant amount of picoplanktonic biomass and productivity in both oceanic and coastal regions.[5] The abundance of Micromonas has increased over the past decade. Evidence shows these spikes in numbers are induced through climate change, which has been felt more drastically in the Arctic.[12] In past years it was thought that green algal species were solely photosynthetic only to discover that was not the case.[25] Micromonas, as well as the Prasinophytes, have taken on mixotrophic lifestyles and have huge impacts on prokaryotic populations within the Arctic.[25] Due to the large consumption of prokaryotes by Micromonas, studies are suggesting photosynthetic picoeukaryotes will soon dominate the primary productivity and bacterivory of Arctic systems.[25] Laboratory studies have shown that species within a single genus have altered their mixotrophic strategies to their distinct environments.[25] These environments can differ through light intensity, nutrient availability and size of prey which all have shown clade-specific alterations to maximize efficiencies.[25]
Viruses are important in the balance of marine ecosystem by regulating the composition of microbial communities, but their behaviors can be affected by several factors including temperature, mode of infection and host conditions.[26][27] There is an increasing number of Micromonas-infecting virus being discovered and studied.
There are currently 45 viral strains identified that coexist with M. pusilla populations.[8] Virus infectivity is dependent on the host strain, light availability and virus adsorption.[28]
Per day average of death due to virus lysis is estimated to be about 2 to 10% of the M. pusilla population.[28]
It is the first phycodnavirus being isolated from polar ocean waters.[31] It can infect M. polaris, which is the polar ecotype of Micromonas that has adapted to waters with low temperatures.[31]
Evidence suggests that the increase in temperature due to climate change may shift the clonal composition of both the virus and host.[31]
With the growing population in the world, there is an increased demand for wild fishes and algae for their source of polyunsaturated fatty acids (PUFA), which is required for growth and development, as well as the maintenance of health in humans. Recent research is investigating an alternative mechanism for production of PUFA by using acyl-CoA Δ6-desaturase, an enzyme present in M. pusilla, with plants. The M. pusilla strain of acyl-CoA Δ6-desaturase is highly effective in the polyunsaturated fatty acid synthesis pathway due to its strong binding preference for omega-3 substrates in land plants.[32]
{{cite book}}: CS1 maint: others (link) {{citation}}: CS1 maint: multiple names: authors list (link) Micromonas is a genus of green algae in the family Mamiellaceae.
Before the characterization in 2016 of a second species, Micromonas commoda, Micromonas pusilla was considered to be the only species in the genus, which led to a disproportionate amount of research discussing a single species within the genus. It is suggested to be the dominant photosynthetic picoeukaryote in some marine ecosystems. Unlike many marine algae, it is distributed widely in both warm and cold waters. It is a strong swimmer and exhibits a phototaxic response.
Micromonas pusilla is divided into 3 to 5 different clades despite their similarity in morphologies and habitats. Varying ratios of clades contribute to the M. pusilla population throughout the marine ecosystem leading to the hypothesis of clades arising based on niche occupation and susceptibility to virus infection.