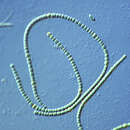
Τα κυανοβακτήρια (Cyanobacteria) αποτελούν φωτοσυνθετικούς οργανισμούς που παλαιότερα κατά την κλασική ταξινόμηση φέρονταν με το όνομα «κυανοφύκη». Σήμερα θεωρούνται ιδιαίτερη συνομοταξία στην επικράτεια των Βακτηρίων. Τα κυανοβακτήρια αποτελούν ομάδα φωτοσυνθετικών προκαρυωτικών οργανισμών, που θεωρούνται ικανά σε οξυγονογενή ή οξυγονούχο φωτοσύνθεση.
Η αρχική τους ονομασία «κυανoφύκη» (blue-green algae στη διεθνή ορολογία) οφειλόταν κυρίως λόγω του χρώματος που παρουσίαζαν πολλά είδη εξ αυτών και που προερχόταν από την παρουσία της βιλιπρωτεΐνης που χαρακτηρίζει τις χρωστικές τόσο των οργανισμών αυτών όσο και των ερυθρών φυκών ή ροδοφύτων, και ειδικότερα της φυκοκυανίνης. Όλα τα μέλη της συνομοταξίας αυτής περιέχουν χλωροφύλλη α, εκτός από τα γένη Πρόχλωρον (Prochloron) και Προχλωροκόκκος (Prochlorococcus) που περιέχουν επίσης και χλωροφύλλη β.
Τα κυανοβακτήρια θεωρούνται ως πιθανοί πρώτοι οργανισμοί στη Γη οι οποίοι παρείχαν οξυγόνο με φωτοσύνθεση. Υπάρχουν αποδείξεις, από απολιθώματα του είδους αυτών, περί της ύπαρξής τους πριν από 3,5 δισεκατομμύρια χρόνια. Το δε κυτταρικό τοίχωμα αυτών είναι ανάλογο με εκείνο των «αρνητικών κατά Gram» βακτηρίων, οι δε φωτεινές αντιδράσεις της φωτοσύνθεσης γίνονται μέσα στο σύστημα της θυλακοειδούς μεμβράνης του κυττάρου. Άλλες δομές του κυττάρου τους είναι διάφορα πολυφωσφορικά σωμάτια που παρατηρούνται σαν απόθεμα φωσφορικών αλάτων, καθώς και τα κενοτόπια που πληρούνται με αέριο και βοηθούν στη πλευστότητα αυτών. Μερικά δε εξ αυτών είναι μονοκύτταρα ενώ άλλα είναι νηματώδη. Αναπαράγονται είτε με ένωση είτε με θραυσματοποίηση.
Τα κυανοβακτήρια βρίσκονται σχεδόν παντού σε ξηρά και θάλασσα και ειδικότερα σε χώρους με άπλετο φωτισμό. Μερικά είδη αυτών ζουν σε τελείως αφιλόξενα περιβάλλοντα μέρη, όπως ακόμη και σε θερμοπηγές όπου η θερμοκρασία υπερβαίνει τους +85 °C. Τα κυανοβακτήρια ευθύνονται σ΄ ένα μεγάλο μέρος της εξέλιξης του φωτοσυνθετικού οξυγόνου ιδιαίτερα στους ωκεανούς και συμβάλουν κατά πολύ στη δέσμευση του διοξειδίου του άνθρακα, κατά τις ανεξάρτητες του φωτός (σκοτεινές) αντιδράσεις καθώς και του αζώτου.
Πολλές φορές συγκεντρώνονται σε μεγάλους αριθμούς κατόπιν υπερβολικής ανάπτυξης σε επιφάνειες λιμνών και υδατοδεξαμενών όπου διακρίνονται τακτικότερα σαν πράσινοι επιπλέοντες κόκκοι. Αυτές όμως οι μεγάλες συγκεντρώσεις μπορεί και να απελευθερώσουν τοξίνες, γνωστές ως κυανοτοξίνες.
Όλοι οι γεωλόγοι και οι γεωχημικοί συμφωνούν ότι τα κυανοβακτήρια έχουν μια μεγάλη εξελικτική ιστορία και οι περισσότεροι, αν όχι όλοι, καταλήγουν στο ότι η ιστορία αυτή επεκτείνεται τουλάχιστον 3500 εκατομμύρια χρόνια πριν. Ωστόσο μια θεωρία σχετικά με την πιθανή ύπαρξη κυανοβακτηρίων στον Άρη προκαλεί ακόμη περισσότερες ερωτήσεις όσον αφορά στην προέλευση τους. Οι Schopf και Walter (1982) έχουν χαρακτηρίσει την πρωτεροζωική περίοδο ως «Εποχή των Κυανοβακτηρίων» , επειδή από εκείνη την περίοδο έχουν βρεθεί άφθονα κυανοβακτήρια σε απολιθωμένη μορφή. Επίσης, μερικές νηματοειδείς δομές που έχουν βρεθεί σε Πρωτοζωικά απολιθώματα έχουν εντυπωσιακή ομοιότητα με τα σύγχρονα νηματοειδή κυανοβακτήρια. Τέλος, ορισμένες δομές, που ήταν κατατεθημένες ευρέως τότε, φαίνεται ότι είναι αρκετά όμοιες με δομές κυανοβακτηρίων που παρουσιάζονται σε περιοχές σήμερα όπως το Shark Bay, στη δυτική Αυστραλία.
Τα κυανοβακτήρια εντοπίζονται σε ένα ευρύ φάσμα περιβαλλόντων. Αυτό αποδίδεται στον ιδιαίτερο μεταβολισμό, που προσδίδει στα κυανοβακτήρια την ικανότητα να μεταβαίνουν γρήγορα από μια λειτουργία σε μια άλλη. Πιο συγκεκριμένα εντοπίζονται σε λίμνες του γλυκού νερού , σε υπεράλμυρα νερά, σε θαλάσσια ύδατα συμπεριλαμβανομένων και των ωκεανών, σε ορυζώνες και ως βενθικοί οργανισμοί σε παράκτια ιζήματα. Επιπλέον, συναντώνται σε εδάφη, ερήμους, σπηλιές και πολλά άλλα περιβάλλοντα. Επίσης, τα κυανοβακτήρια είναι γνωστά ως συμβιωτικοί οργανισμοί, όπως για παράδειγμα στο θαλάσσιο διάτομο Rhizosolenia, στις ρίζες του γένους Cycas και στα φύλλα του γένους Azolla.
Οι συμβιωτικές σχέσεις των κυανοβακτηρίων με άλλους οργανισμούς αποτελούν μια από τις αρχαιότερες αλληλεπιδράσεις και είναι ιδιαίτερα διαδεδομένες τόσο σε υδρόβια όσο και σε χερσαία περιβάλλοντα. Στους οργανισμούς με τους οποίους συμβιούν τα κυανοβακτήρια περιλαμβάνονται μύκητες, φυτά και ζώα. Στην πλειονότητα αυτών των σχέσεων, τα κυανοβακτήρια παρέχουν στον ξενιστή άζωτο μέσω της ικανότητας τους για αζωτοδέσμευση, επιτρέποντας του να ζει σε περιβάλλοντα φτωχά σε άζωτο. Παράλληλα, τα κυανοβακτήρια επιτρέπουν στους μη φωτοσυνθετικούς ξενιστές τους να εκμεταλλεύονται την ηλιακή ενέργεια, καλύπτοντας μέρος των ενεργειακών τους αναγκών, μέσω της φωτοσυνθετικής τους δραστηριότητας. Ακόμη, προσφέρουν προστασία από την υπεριώδη ακτινοβολία και παράγουν μια ποικιλία βιοδραστικών ουσιών (αντιβιοτικά και αντιοξειδωτικά) συμμετέχοντας στη χημική άμυνα και προστασία του ξενιστή. Τα οφέλη που λαμβάνουν τα κυανοβακτήρια από τους ξενιστές τους περιλαμβάνουν ένα σταθερό υπόστρωμα, προστατευμένο από τις ακραίες περιβαλλοντικές συνθήκες και τη θήρευση. Σε ορισμένες περιπτώσεις, ο ξενιστής μπορεί να παρέχει άνθρακα στα κυανοβακτήρια που φιλοξενεί, τα περισσότερα από τα οποία αν και είναι φωτοαυτότροφοι οργανισμοί, μπορούν να ζήσουν και σαν προαιρετικά ετερότροφοι, γεγονός που τους δίνει τη δυνατότητα να ζουν σε περιοχές του ξενιστή όπως οι ρίζες των φυτών όπου δεν φτάνει το φως. Σε αυτή την περίπτωση, ο φωτοσυνθετικός ξενιστής τους καλύπτει εξολοκλήρου τις απαιτήσεις τους σε άνθρακα. Πολλές από τις συμβιωτικές σχέσεις που σχηματίζουν τα κυανοβακτήρια, αποτελούν σημαντικές πηγές αζώτου για το οικοσύστημα. Συμβιωτικά κυανοβακτήρια έχουν εντοπιστεί στις παρακάτω κατηγορίες οργανισμών:
Τέλος, συμβιωτικά κυανοβακτήρια έχουν καταγραφεί σε μακροφύκη, τρηματοφόρα, ακτινόζωα, κωδωνοειδή, ευγλείνες και ισόποδα.
Οι μαζικές συναθροίσεις των κυανοβακτηρίων είναι γνωστές με τον όρο «ανθίσεις του νερού» water blooms και χαρακτηρίζονται τόσο από πυκνό αριθμό κυττάρων διασκορπισμένων στο νερό όσο και από το σχηματισμό στρωμάτων ακαθαρσιών στην επιφάνεια. Ορισμένα κυανοβακτήρια παράγουν κατά τη φυσιολογική τους ανάπτυξη χαρακτηριστικές οσμές. Η παρουσία των οσμηρών ουσιών, ωστόσο, δε σχετίζεται απαραίτητα με τοξικότητα και μπορεί να λαμβάνονται ως ένα προειδοποιητικό σήμα για την ύπαρξη των κυανοβακτηρίων. Ενώ οι συναθροίσεις των κυανοβακτηρίων προκαλούνται συνήθως από πλαγκτικά είδη σε ευτροφικά ύδατα, βενθικά στρώματα σε ολιγοτροφικά νερά είναι επίσης δυνατό να προκαλέσουν προβλήματα. Οι επιβλαβείς ανθήσεις των κυανοβακτηρίων ονομάζονται CyanoHABs και αποτελούν έναν από τους πιο εμφανείς μικροβιακούς κινδύνους για την ποιότητα των υδάτων των γλυκών και θαλάσσιων οικοσυστημάτων, την παροχή πόσιμου νερού, την άρδευση, την αλιεία και την αναψυχή. Ο κίνδυνος αυτός προκύπτει από την παραγωγή επιβλαβών δευτερογενών μεταβολιτών που ονομάζονται κυανοτοξίνες και περιλαμβάνουν τις μικροκυστίνες, σαξιτοξίνες κτλ [4][5]. Εμβαθύνοντας, έρευνες δείχνουν ότι, οι αστικές, οι γεωργικές και οι βιομηχανικές δραστηριότητες που σχετίζονται με τον αυξημένο παγκόσμιο ανθρώπινο πληθυσμό, έχουν προωθήσει την ανακατανομή των θρεπτικών συστατικών. Το γεγονός αυτό οδηγεί σε φαινόμενα ευτροφισμού τα οποία διεγείρουν τα CyanoHABs[6].
Ενώ η σύνδεση μεταξύ του ευτροφισμού και της αύξησης της συχνότητας και της έντασης των CyanoHABs είναι καλά εδραιωμένη, έρευνες που διεξάγονται σχετικά με την οικοφυσιολογίας και της δυναμικής των κυανοβακτηρίων υποδηλώνει ότι, θα επηρεαστούν θετικά υπό τις συνθήκες που δημιουργούνται από την παγκόσμια κλιματική αλλαγή. Πιο συγκεκριμένα, η υπερθέρμανση του πλανήτη και οι ταυτόχρονες υδρολογικές μεταβολές, μεταβάλουν έντονα το φυσικο-χημικό περιβάλλον και τις βιολογικές διεργασίες των κυανοβακτηρίων, παρέχοντας στα κυανοβακτήρια ένα ξεχωριστό πλεονέκτημα έναντι του ευκαρυωτικού φυτοπλαγκτού, υπό συνθήκες εμπλουτισμένες με θρεπτικά συστατικά. Αυτό διότι, οι ρυθμοί ανάπτυξης των κυανοβακτηρίων φθάνουν στο βέλτιστο επίπεδο και παραμένουν υψηλοί ακόμη και όταν οι θερμοκρασίες υπερβαίνουν τους 25 ° C (όπως σε στελέχη του γένους Microcystis) ενώ, οι ρυθμοί ανάπτυξης των ευκαρυωτικών taxa σταθεροποιούνται ή μειώνονται[7][5][6]. Επίσης, η αύξηση της θερμοκρασίας σε μεγαλύτερα γεωγραφικά πλάτη, έχει προκαλέσει την εξάπλωση τοξικών κυανοβακτηρίων, όπως το Cylindrospermopsis raciborskii σε περιοχές, όπου δεν υπήρχαν προηγουμένως καταγραφές[8]. Ακόμα, η θέρμανση των επιφανειακών υδάτων σε συνδυασμό με την εποχική διακύμανση, επιτυγχάνει και εντείνει την κατακόρυφη στρωμάτωση των λιμναίων κυρίως υδάτων. Στη συγκεκριμένη περίπτωση, δημιουργούνται οι κατάλληλες προϋποθέσεις για την άνθηση πολλών ειδών κυανοβακτηρίων που περιέχουν αεροτόπια, παρέχοντάς τους πλευστότητα και πρόσβαση στα επιφανειακά στρώματα υδάτων με τα υψηλά επίπεδα ακτινοβολίας για αποδοτικότερη φωτοσύνθεση. Αυτά τα κυανοβακτήρια, περιέχουν φωτοπροστατευτικές βοηθητικές χρωστικές ουσίες (π.χ. καροτενοειδή) και ενώσεις απορρόφησης υπεριώδους ακτινοβολίας που εξασφαλίζουν μακροπρόθεσμη επιβίωση υπό εξαιρετικά υψηλές συνθήκες ακτινοβολίας, καταστέλλοντας τα υπόλοιπα είδη μέσω ανταγωνισμού για το φως. Παρατηρείται, λοιπόν, ότι τα κυανοβακτήρια έχουν αποτελεσματικούς μηχανισμούς πρόσληψης CO2 και θρεπτικών συστατικών, προστατεύονται καλά από την ακτινοβολία του φωτός και της υπεριώδους ακτινοβολίας και είναι ιδιαίτερα προσαρμόσιμοι, επιτρέποντάς τους να επωφεληθούν από τις μεταβαλλόμενες περιβαλλοντικές συνθήκες σε υδάτινα περιβάλλοντα[6]. Επιπροσθέτως, στα αποτελέσματα της κλιματικής αλλαγής συγκαταλέγονται και οι κλιματικές ταλαντώσεις οι οποίες επηρεάζουν τα πρότυπα, τις εντάσεις και τη διάρκεια των βροχοπτώσεων και των ξηρασιών έχοντας αντίκτυπο στην κατανομή των CyanoHABs. Για παράδειγμα, πιο ακραία γεγονότα κατακρημνίσεων μεταφέρουν μεγαλύτερο φορτίο θρεπτικών συστατικών από την χέρσο στα υδάτινα οικοσυστήματα-υποδοχείς. Αυτό το σενάριο πιθανόν θα συμβεί αν οι αυξημένες βροχοπτώσεις χειμώνα-άνοιξη και τα γεγονότα εξάτμισης ακολουθούνται από παρατεταμένες περιόδους ξηρασίας. Παρόλα αυτά η δύναται να αυξηθεί η εκροή των γλυκών υδάτων στα παρακείμενα ύδατα και βραχυπρόθεσμα θα κατασταλεί η άνθηση με έξαψη[6]. Επιπλέον, ενδεχομένως η κλιματική αλλαγή μπορεί επίσης να επηρεάσει την αλατότητα στις εκβολές των ποταμών και των λιμών λόγω της αύξησης της στάθμης της θάλασσας (αύξηση της συχνότητας και της διάρκειας της ξηρασίας με συνακόλουθη την αύξηση της αποξήρανσης ή αύξηση των βροχοπτώσεων λόγω καταιγίδων). Ως αποτέλεσμα, τροποποιούνται τόσο η σύνθεση του φυτοπλαγκτού, όσο και οι πιθανές συγκεντρώσεις και η κατανομή τοξινών διότι, αν και πολλά ευκαρυωτικά είδη φυτοπλαγκτού δεν μπορούν να ανεχθούν μεταβολές στην αλατότητα, ορισμένα είδη κυανοβακτηρίων εμφανίζουν μεγάλη ανοχή σ’ αυτές. Είναι γνωστό ότι ένας βασικός παράγοντας που συντελεί στην κλιματική αλλαγή αποτελεί η αύξηση των επιπέδων του ατμοσφαιρικού διοξειδίου του άνθρακα (CO2) που συνδέεται με το φαινόμενο του θερμοκηπίου. Γενικά, αν και οι λεπτομέρειες για το πώς συγκεκριμένα γένη κυανοβακτηρίων μπορεί να ανταποκριθούν στην αλλαγή του κλίματος δεν είναι πλήρως κατανοητές, στα είδη που σχηματίζουν επιπολασμό ανήκουν κυρίως στα γένη Microcystis, Anabaena και Aphanizomenon[7] .
Η ενδεχόμενη παρουσία κυανοβακτηρίων σε υπόγειους υδροφόρους ορίζοντες, μπορεί να αποτελέσει σημαντικό πρόβλημα, τόσο για τη δημόσια υγεία, όσο και για το περιβάλλον. Μέσω των υπογείων υδάτινων όγκων, τα κυανοβακτήρια μπορούν να διοχετευθούν τόσο στο πόσιμο νερό όσο και στα θαλάσσια οικοσυστήματα. Ο κίνδυνος προέρχεται από συγκεκριμένα είδη δυνητικά τοξικών κυανοβακτηρίων, τα οποία παράγουν κυανοτοξίνες, δηλαδή δευτερογενείς μεταβολίτες που έχουν επιβλαβείς επιδράσεις, όταν βρίσκονται είτε σε κλειστές δεξαμενές νερού, είτε σε ρέοντα ύδατα.
Οι επιβλαβείς πληθυσμιακές εκρήξεις βακτηρίων έχουν αυξηθεί τα τελευταία χρόνια [9]. Στην Ελλάδα, το θερμό Μεσογειακό κλίμα, ευνοεί την ανάπτυξη κυανοβακτηρίων, ιδίως τους θερμούς μήνες του χρόνου. Η αύξηση της θερμοκρασίας του πλανήτη συντελεί στη δημιουργία ιδανικών συνθηκών για απότομες πληθυσμιακές αυξήσεις δυνητικά τοξικών κυανοβακτηρίων, καθώς και την επικράτηση των τοξικών έναντι των μη τοξικών στελεχών.
Το εμφιαλωμένο νερό μπορεί να προέρχεται από φυσικούς υδροφόρους ορίζοντες, πηγές, λιώσιμο παγετώνων καθώς και δημόσιες προμήθειες νερού. Σε κάθε περίπτωση υπάρχει ο κίνδυνος μόλυνσης με μικροκυστίνες και απαιτείται συνεχής παρακολούθηση των υδάτων για τον εντοπισμό τους. Για να χαρακτηριστεί το εμφιαλωμένο νερό ως φυσικό μεταλλικό, η μόνη επεξεργασία που επιτρέπεται να υποστεί είναι το φιλτράρισμα, το οποίο, αν και συγκρατεί το μεγαλύτερο ποσοστό των μικροφυκών, μπορεί να επιτρέπει σε πιθανές τοξίνες που έχουν παραχθεί από αυτά, να περάσουν στο τελικό προϊόν. Στην Ελλάδα δεν υπάρχει νομοθεσία για την παρακολούθηση των υπόγειων υδάτων. Ωστόσο, μια έρευνα που πραγματοποιήθηκε στην Ελλάδα και περιλάμβανε τον δειγματικό έλεγχο τόσο εμφιαλωμένου νερού όσο και υπογείων υδάτων για κυανοβακτήρια, εντόπισε μια αποικία σε ένα εκ των δειγμάτων εμφιαλωμένου νερού. Η πηγή της μόλυνσης ωστόσο δεν εντοπίστηκε, επομένως, το ενδεχόμενο επιμόλυνσης μέσω του αέρα, κατά τη διαδικασία της εμφιάλωσης, δεν μπορεί να αποκλειστεί.
Το εύρημα αυτό, αν και δεν είναι αρκετό για την απόδειξη άμεσου κινδύνου, αλλά εγείρει ερωτήματα για το αν θα πρέπει να νομοθετηθεί πρόγραμμα ελέγχου των υπόγειων υδάτων.
Έχουν γνωστοποιηθεί περίπου 2000 είδη κυανοβακτηρίων τα οποία μπορούν να χωριστούν σε 150 γένη. Επειδή πολλά κυανοβακτήρια στερούνται ευδιάκριτων μορφολογικών χαρακτηριστικών, η ταξινόμηση τους είναι ιδιαίτερα δύσκολη και διαρκώς αναδιατάσσεται. Οι σύγχρονες οικολογικές μελέτες, η ηλεκτρονική μικροσκοπία, καθώς και οι μοριακές τεχνικές έχουν επηρεάσει τη γνώση σχετικά με την ποικιλομορφία των κυανοβακτηρίων. Στο εγχειρίδιο Bergey’s Manual of systematic Bacteriology η ταξινόμηση των κυανοβακτηρίων γίνεται σε πέντε υποκατηγορίες, οι οποίες αντιστοιχούν στην ταξινομική ομάδα Τάξη και είναι οι ακόλουθες : α)Τάξη Chroococcales , στην οποία συμπεριλαμβάνονται μονοκύτταρα κυανοβακτήρια που αναπαράγονται με απλή διχοτόμηση, β)Τάξη Pleurocapsales , στην οποία εμπεριέχονται μονοκύτταρα κυανοβακτήρια που αναπαράγονται με πολλαπλή διχοτόμηση, γ)Τάξη Oscillatoriales , δ) Τάξη Nostocales και ε) Τάξη Stigonematales. Οι τρεις τελευταίες Τάξεις περιέχουν τα νηματοειδή κυανοβακτήρια.
Τα τελευταία χρόνια τα κυανοβακτήρια θεωρούνται σημαντικά ως πλούσια πηγή βιοδραστικών συστατικών. Αυτά τα συστατικά περιλαμβάνουν αντιβακτηριακούς ,αντιμυκητιασικούς , αντι-ιικούς , αντικαρκινικούς και ανοσοκατασταλτικούς παράγοντες. Έχει βρεθεί ότι μερικά κυανοβακτήρια συσσωρεύουν ενδοκυτταρικά PHA’s (polyhydroxyalkanoates) , που αποτελούν πολυεστέρες με συγκρίσιμες ιδιότητες σε αυτές του πολυαιθυλενίου και του πολυπροπυλενίου. Αυτά τα βιοδιασπώμενα πλαστικά θα μπορούσαν να αντικαταστήσουν σε ορισμένα πεδία τα θερμοπλαστικά που είναι παράγωγα του πετρελαίου. Πρόσφατη έρευνα κατέδειξε ότι τα κυανοβακτήρια είναι πιθανό να χρησιμοποιηθούν για τον καθαρισμό ιζημάτων και λυμάτων που έχουν μολυνθεί με πετρέλαιο. Επιπλέον , παρουσιάζουν κι άλλες βιοτεχνολογικές εφαρμογές που αφορούν σε πιθανές χρήσεις σε υδατοκαλλιέργειες , τρόφιμα, καύσιμα, λιπάσματα, χρωστικές ουσίες και στην παραγωγή δευτερογενών μεταβολιτών συμπεριλαμβανομένων τοξινών, βιταμινών, ενζύμων και φαρμακευτικών παρασκευασμάτων.
Τα κυανοβακτήρια έχουν την ικανότητα παραγωγής βιομορίων που διεγείρουν το εμπορικό και επιστημονικό ενδιαφέρον του ανθρώπου. Η ικανότητα αυτή οφείλεται κυρίως στην δυνατότητα που έχουν να μετατρέπουν ανόργανα στοιχεία, όπως για παράδειγμα διοξείδιο του άνθρακα (CO2), νιτρικά και φωσφορικά ιόντα ή ιχνοστοιχεία σε βιομάζα και οργανικούς μεταβολίτες[10]. Έχει αναφερθεί ότι στο δυναμικό παραγωγής τους συμπεριλαμβάνονται περίπου 1100 μεταβολίτες του δευτερογενούς μεταβολισμού όπως τερπένια, καροτενοειδή, αλκαλοειδή, λιπίδια με αντικαρκινικές, αντιμικροβιακές, αντιικές ιδιότητες κ.α. Εκτός από την καλλιέργεια φυσικών στελεχών και έπειτα την συγκομιδή της βιομάζας για περαιτέρω αξιοποίηση, καλλιεργούνται ήδη και γενετικά τροποποιημένα στελέχη για μαζική παραγωγή βιοκαυσίμων, τερπερνοειδών, λιπαρών οξέων και σακχάρων. Πιο συγκεκριμένα, τα στελέχη Synechocystis sp. PCC 6803, Synechococcus sp. PCC 7002 και Synechococcus elongatus PCC 7942 αποτελούν αντικείμενα μελέτης αρκετών πειραμάτων γενετικής μηχανικής με σκοπό μαζικότερη και ποιοτικότερη παραγωγή βιοπροϊόντων. Επιπλέον, χρησιμοποιούνται ευρέως ως «οργανισμοί-μοντέλα» για τη διερεύνηση της φωτοσύνθεσης, της αζωτοδέσμευσης και άλλων μεταβολικών μονοπατιών με πιθανή βιομηχανική χρηστικότητα.
Η βελτίωση κάθε σταδίου της βιομηχανικής παραγωγής επιτυγχάνεται χάρη στην πληθώρα γενετικών εργαλείων που προσφέρει ο κλάδος της συνθετικής βιολογίας. Ο κλάδος αυτός συνδέει την γενετική μηχανική με τον σχεδιασμό πρωτεϊνικών δομών μέσω υπολογιστικών προγραμμάτων μοντελοποίησης. Το γεγονός αυτό επιτυγχάνεται με τον σχεδιασμό βιολογικών τμημάτων (π.χ. υποκινητές, τερματιστές κ.λπ.) που επιτελούν το κάθε ένα μια καθορισμένη λειτουργία, και που αν συγκροτηθούν μπορούν να σχηματίσουν ένα εργαλείο με καθορισμένη δράση. Το συνθετικό τμήμα εισάγεται στο γονιδίωμα με φορείς που επιτρέπουν την ενσωμάτωση του μέσω ομόλογου ανασυνδυασμού. Με αυτό τον τρόπο κατασκευάζονται εργαλειοθήκες με σκοπό τον έλεγχο της γονιδιακής έκφρασης. Σχετικά με την πρόοδο της συνθετικής βιολογίας στα κυανοβακτήρια, οι Markley et al. (2015) αναφέρουν την κατασκευή δύο νέων υποκινητών βασιζόμενη στον ιδιοστατικό υποκινητή cpcB που υπάρχει στο Synechocystis sp. PCC 6803 [11]. Παρόλα αυτά, η δημιουργία γενετικών εργαλείων που να εφαρμόζονται σε γονιδιώματα κυανοβακτηρίων αποτελεί πρόκληση λόγω της μεγάλης ενδοειδικής ποικιλότητας που εμφανίζουν. Ακόμη, σημαντικά είναι τα αποτελέσματα πειραματικών προσπαθειών που αφορούν το μεταβολισμό του άνθρακα και καταδεικνύουν ότι η υπερέκφραση των γονιδίων που κωδικοποιούν ένζυμα που εμπλέκονται στον κύκλο, οδηγούν σε αυξημένα επίπεδα φωτοσυνθετικής δραστηριότητας (συνεπώς και παραγωγής βιομάζας) στα υπό μελέτη κυανοβακτήρια. Mε την ίδια λογική πραγματοποιήθηκαν προσπάθειες υπερέκφρασης του ενζύμου RuBisCO (ριβουλόζη-1,5-διφωσφορική καρβοξυλάση/οξυγενάση) που διαδραματίζει σημαντικό ρόλο στο μεταβολισμό των κυανοβακτηρίων με στόχο την αύξηση των επιπέδων δέσμευσης CO2 και συνεπώς την αύξηση της αποδοτικότητας. Παρόλα αυτά, η μέχρι στιγμής έρευνα υποδεικνύει τη διεξαγωγή επιπλέον μελετών που αφορούν πρωτίστως την κατανόηση της έκφρασης των γονιδίων στο οπερόνιο rbc και τον τρόπο συναρμολόγησης της ενζυματικής συσκευής, εξαιτίας της πολύπλοκης γονιδιακής ρύθμισης στην οποία υπόκειται.
Τα προϊόντα που προκύπτουν από μεθόδους συνθετικής βιολογίας διακρίνονται σε υψηλής, μέτριας και χαμηλής αξίας. Λόγω της σχετικής ευκολίας των κυανοβακτηρίων να τροποποιούνται σε γενετικό επίπεδο, θεωρούνται η βασική πηγή για την παραγωγή βιοκαυσίμων 4ης γενιάς. Ακόμη, υπό υλοποίηση σε εργαστηριακό περιβάλλον (Proterro, J. Craig Venter Institute κ.ά.) βρίσκεται η δημιουργία γενετικά τροποποιημένων κυανοβακτηρίων με ενισχυμένο μεταβολισμό για απευθείας παραγωγή και απελευθέρωση υδρογόνου, αλκοολών και αναλόγων πετρελαίου με στόχο την αποφυγή κοστοβόρων διαδικασιών που περιλαμβάνει η συγκομιδή της βιομάζας και η επεξεργασία της. Αύξηση στην παραγωγή αιθανόλης σε τίτλους έως και 5.5 g/L χρησιμοποιώντας μεθόδους συνθετικής βιολογίας παρατηρήθηκε από τους Gao et al. (2012) και μέσω του σχεδίου DEMA (Direct Ethanol from Microalgae) δρομολογούνται περαιτέρω μελέτες για μείωση του κόστους παραγωγής ενώ διερευνάται το ενδεχόμενο παραγωγής και άλλων μεγαλύτερης αξίας αλκοολών από στελέχη Synechococcus (1-βουτανόλη, ισοπροπανόλη, βουτανοδιόλη)[12]. Επιπλέον, επιτυχημένες πειραματικές προσπάθειες έχουν διεξαχθεί σε κύτταρα του στελέχους Synechocystis sp. PCC 6803 με αποτέλεσμα την μεγιστοποίηση παραγωγής ελεύθερων λιπαρών οξέων ενώ με τη σύνθεση και ενσωμάτωση ενός συστήματος κυτταρικής λύσης από τους Liu & Curtis (2009) κατέστη δυνατή η απελευθέρωση συσσωρευμένων λιπιδίων εντός του κύτταρου[13]. Τέλος, σχετικά με το ενδεχόμενο αύξησης του ποσού υδρογόνου που μπορεί να παραχθεί από νηματοειδή κυανοβακτήρια με ετεροκύτια, η προσέγγιση που προτείνει η συνθετική βιολογία περιλαμβάνει την ανάπτυξη συστημάτων που εμπεριέχουν την εισαγωγή στα κυανοβακτήρια αλληλουχιών υδρογενασών άλλων μικροοργανισμών. Τέλος, έντονη ερευνητική και βιομηχανική δραστηριότητα παρατηρείται και στη βελτίωση παραγωγής τερπενίων για φαρμακευτικούς κυρίως σκοπούς.
Η γενετική τροποποίηση κυανοβακτηριακών στελεχών με μεθόδους συνθετικής βιολογίας, ενέχει τον κίνδυνο διαρροής τροποποιημένων κυττάρων στο φυσικό περιβάλλον σε κάποιο από τα στάδια της καλλιέργειας ή των πειραματικών διεργασιών. Ένα τέτοιο γεγονός θα μπορούσε να προκαλέσει την αποσταθεροποίηση του εκάστοτε οικοσυστήματος, ευνοώντας την επικράτηση των τροποποιημένων στελεχών εις βάρος των φυσικών πληθυσμών και αλληλεπιδρώντας μαζί τους μέσω οριζόντιας μεταφοράς γονιδίων. Για να αποφευχθούν αντίστοιχες απειλές της ποικιλότητας του φυτοπλαγκτού στο φυσικό περιβάλλον, έχουν αναπτυχθεί μηχανισμοί - διακόπτες ικανοί να ενσωματώνονται στο γονιδίωμα των τροποποιημένων στελεχών με στόχο να προκαλούν προγραμματισμένο κυτταρικό θάνατο σε συνθήκες εκτός καλλιέργειας. Πιο συγκεκριμένα, οι μηχανισμοί αυτοί έχουν την δυνατότητα να προκαλούν το θάνατο του τροποποιημένου κυττάρου είτε ενεργά, με τη δράση πρωτεϊνών που αποδιατάσουν το RNA ή καταστρέφουν το DNA, την κυτταρική μεμβράνη ή το κυτταρικό τοίχωμα, είτε παθητικά, με την βιωσιμότητα των συνθετικά τροποποιημένων κυττάρων να εξαρτάται από κάποια χημική ένωση που παρέχεται στο μέσο καλλιέργειας αλλά είναι σπάνια στο φυσικό περιβάλλον.
Τα κυανοβακτήρια παράγουν ποικίλες βιολογικά και χημικά δραστικές ενώσεις αρκετές από τις οποίες έχουν μελετηθεί εκτενώς. Αυτοί οι μεταβολίτες μπορεί να παρουσιάζουν ηπατοτοξικές, νευροτοξικές ή κυτταροτοξικές ιδιότητες και συνήθως είναι κυκλικά πεπτίδια, λιποπεπτίδια, λιπαρά οξέα, αλκαλοειδή και σακχαρίτες. Η βιοσύνθεσή τους πραγματοποιείται κυρίως κατά την άνθιση των κυανοβακτηρίων και είναι συνήθως πεπτίδια που δε συντίθενται στα ριβοσώματα. Παρόλα αυτά ο μηχανισμός βιοσύνθεσης πολλών παραμένει άγνωστος. Οι ενεργότητες που μπορεί να εμφανίζουν οι τοξικοί μεταβολίτες είναι: παρέμβαση στη μεταγωγή σήματος είτε ενεργοποιώντας ή αποφράσσοντας κανάλια νατρίου είτε στοχεύοντας πρωτεΐνες σηματοδότησης, διάσπαση πρωτεϊνών του κυτταροσκελετού επάγοντας έτσι απόπτωση, αναστολή μεμβρανικών υποδοχέων, μεταφορέων, πρωτεασών σερίνης κλπ. Οι μεταβολίτες των θαλάσσιων κυανοβακτηρίων παρουσιάζουν ενδιαφέρον στη φαρμακολογία λόγω των ιδιοτήτων τους να ελέγχουν την ανάπτυξη καρκινικών κυτταρικών σειρών και παθογόνων μικροοργανισμών (βακτήρια, ιοί, μύκητες, πρωτόζωα)
Παρόλο που έχουν αναγνωριστεί ποικίλες κατηγορίες δραστικών προιόντων των κυανοβακτηρίων τα πεπτίδια επικρατούν. Σημαντικός αντιπρόσωπος στα θαλάσσια κυανοβακτήρια είναι οι απρατοξίνες ο σκελετός των οποίων αποτελείται από πεπτίδια και πολυκετίδια. Συγκεκριμένα η απρατοξίνη Α που απομονώθηκε από το Lyngbya majuscula εμφάνισε σημαντική κυτταροτοξικότητα σε κυτταρικές σειρές ανθρώπινων όγκων (in vitro) και επέφερε πλήρη ανάρρωση σε ποντίκια που βρίσκονταν σε πρώιμο στάδιο αδενοκαρκινώματος (in vivo). Η αντινεοπλασματική ιδιότητα της απρατοξίνης Α αποδίδεται στην ικανότητά της να σταματά τον κυτταρικό κύκλο στη φάση G1 σε κύτταρα όγκου. Επιπλέον εμποδίζει την ενεργοποίηση και φωσφορυλίωση της ογκογόνου πρωτεΐνης STAT3 (signal transducer and activator of transcription 3). Τέλος μπορεί να ρυθμίσει αρνητικά μερικούς υποδοχείς κινάσης τυροσίνης (RTKs) με τελικό αποτέλεσμα την αποικοδόμησή τους από το πρωτεάσωμα. Άλλες απρατοξίνες είναι οι Β με ασθενέστερη κυτταροτοξικότητα συγκριτικά με την Α, η C με ισχυρή κυτταροτοξικότητα, οι D,E,F και G.
Οι μικροσωληνίσκοι αποτελούνται από διμερή τουμπουλίνης και νηματίων ακτίνης. Σχηματίζουν τη μιτωτική άτρακτο που είναι απαραίτητη για το διαχωρισμό των αδελφών χρωματίδων κατά τη μίτωση. Οποιαδήποτε παρέμβαση στο μηχανισμό αυτό οδηγεί σε διακοπή της μίτωσης και πιθανώς απόπτωση. Στους παράγοντες που διασπούν τους μικροσωληνίσκους άρα ελέγχουν τον πολλαπλασιασμό καρκινικών κυττάρων και προέρχονται από θαλάσσια κυανοβακτήρια ανήκουν τα πεπτίδια δολαστατίνης τα οποία έχουν την ιδιότητα να αναστέλλουν τη συναρμολόγηση των μικροσωληνίσκων διαλύοντας έτσι την μιτωτική άτρακτο.
Τα κανάλια Nα+ και Ca+2 που βρίσκονται στις κυτταρικές μεμβράνες συμμετέχουν ενεργά στη μεταγωγή σημάτων εντός και εκτός του κυττάρου. Πολλές έρευνες απέδειξαν ότι νευροτοξίνες που παράγονται από διάφορα είδη κυανοβακτηρίων εμπλέκονται τόσο στην ενεργοποίηση όσο και στην αναστολή των καναλιών της μεμβράνης επηρεάζοντας την μεταγωγή σήματος. Ενδεικτικά, η χοιαμίδη Α που απομονώθηκε από τα είδη L. majuscula και Phormidium gracile ενεργοποιεί τα κανάλια Να+ ενώ η αντιλατοξίνη και η κακλιτοξίνη προκαλούν νευρική νέκρωση. Επίσης, τα είδη των κυανοβακτηρίων Anabaena circinalis, A. perturbata var. tumida και Α. spiroides παράγουν την σαξιτοξίνη που αναστέλει τα κανάλια Να+, Ca+2 και Κ+ ενώ η Μ1 αντιτοξίνη από το είδος Nostoc sp. εμποδίζει την προσβολή την ηπατικών κυττάρων απο ηπατοτοξίνες αναστέλλοντας την λειτουργία ενός μεταφορέα ανιόντων. Όλες αυτές οι λειτουργίες καθιστούν τα κυανοβακτήρια σημαντική πηγή ουσιών για την παραγωγή αναισθητικών, αναλγητικών, σπασμολυτικών και αντιεπιληπτικών φαρμάκων.
Πολλά είδη βενθικών και πλαγκτικών κυανοβακτηρίων παράγουν μεταβολίτες που έχουν την ικανότητα να σταματούν την ανάπτυξη των παθογόνων μικροοργανισμών. Το είδος L. majuscula παράγει τα ολιγοπεπτίδια καρμαμπίνη Α, δραγομαβίνη και δραγοναμίδη Α τα οποία δρουν αποτελεσματικά κατά της ελονοσίας μέσω της αναστολής των ανθεκτικών στην κινίνη στελεχών. Παρόμοια είναι η δράση της βεντουραμίδης Α και Β που απομονώθηκαν από το είδος Oscillatoria sp. Το L. majuscula παράγει, επίσης, τη δραγοναμίδη Ε που εμποδίζει την ανάπτυξη του παθογόνου πρωτοζώου Leishmania . Οι βιρινταμίδες Α και Β από το είδος Oscillatoria nigro-viridis εμποδίζουν την ανάπτυξη τόσο του Leishmania όσο και του τρυπανοσώματος ενώ το είδος Lyngbya confervoides παράγει τις λοβοκυκλαμίδες Α-C από τις οποίες οι Α και Β έχουν αντιμυκητιακή δράση. Όλοι αυτοί οι μεταβολίτες δρουν επιλεκτικά ενάντια σε παθογόνους μικροοργανισμούς και παράσιτα διακόπτοντας τα μεταβολικά μονοπάτια των οργανισμών που στοχεύουν.
Είναι γνωστό ότι τα κυανοβακτήρια μπορούν να παράγουν επικίνδυνα μόρια για την ανθρώπινη υγεία, η παραγωγή όμως των γνωστών κυανοτοξίνων είναι ταξινομικά σποραδική. Για παράδειγμα κάποια είδη παράγουν μικροκυστίνες που είναι αρκετά επιβλαβείς στην ανθρώπινη υγεία, Μια νευροτοξίνη β-Ν-μεθυλαμινο-L-αλανίνη (ΒΜΑΑ), φαίνεται να παράγεται από όλες τις γνωστές ομάδες κυανοβακτηρίων συμπεριλαμβανομένων των συμβιωτικών και ελεύθερων. Ο άνθρωπος είναι δυνατό να έρθει σε επαφή με τα κυανοβακτήρια από αρκετές πηγές όπως το νερό και το έδαφος.
Το β-Ν-μεθυλαμινο-L-αλανίνη (BMAA),είναι ένα μη πρωτεϊνικό αμινοξύ που βρέθηκε ότι παράγεται από συμβιωτικά κυανοβακτήρια στις ρίζες του γένους Nostoc. Αρχικά ανακαλύφθηκε σε ένα είδος φοινικόθαμνου και είναι υπεύθυνο για αρκέτες παθήσεις. Επιπρόσθετα έχει βρεθεί ότι το ΒΜΑΑ μπορεί να ενσωματωθεί λανθασμένα αντί της σερίνης στις ανθρώπινες πρωτεΐνες και έτσι αυξάνεται η πιθανότητα εμφάνισης νευροεκφυλιστηκών ασθενειών. Το BMAA προτάθηκε ακόμα να είναι υπεύθυνο για την αμυοτροφική πλευρική σκλήρυνση και τον παρκινσονισμό. Επίσης, ανακαλύφθηκε πρόσφατα σε 9 ασθενείς στον Καναδά που έπασχαν από την ασθένεια του Αλτσχάιμερ. Βρέθηκε ακόμα ότι η λανθασμένη αυτή τοποθέτηση είναι υπεύθυνη για την άνοια και την προϊούσα υπερπυρηνική παράλυση.
Μια πρόσφατη μελέτη αναφέρει ότι το BMAA συσσωρεύεται σε πρωτεΐνες , το όποιο επιλεκτικά λειτουργεί σαν ενδογενής δεξαμενή νευροτοξίνης μέσα στο ανθρώπινο σώμα και απελευθερώνεται σταδιακά με το χρόνο και οι πρωτεΐνες αυτές μεταβολίζονται.
Τέλος, το ΒΜΑΑ μπορεί να πυροδοτήσει νευροεκφυλισμό σε ευάλωτα άτομα. Για την αποφυγή αυτού του νευροεκφυλισμού μια λύση που μελετάται είναι η αυξημένη πρόσληψη σερίνης στο καθημερινό διαιτολόγιο.
Η ιστορία των φυσικών επιστημών σχετίζεται άμεσα με τον ελληνικό πολιτισμό, αφού, οι παλαιότερες ακόμα γραπτές αναφορές σε φυτά, ζώα, και φύκη βρίσκονται σε Ομηρικά έπη (8ος αιώνας π.Χ.) Η πρώτη αναφορά σε φύκη στη δυτική λογοτεχνία εμφανίζεται στην Ιλιάδα: [Ραψωδία ι, στ. 5-9]
«Βορέης καὶ Ζέφυρος, τώ τε Θρῄκηθεν ἄητον ἐλθόντ᾽ ἐξαπίνης· ἄμυδις δέ τε κῦμα κελαινὸν κορθύεται, πολλὸν δὲ παρὲξ ἅλα φῦκος ἔχευεν· ὣς ἐδαΐζετο θυμὸς ἐνὶ στήθεσσιν Ἀχαιῶν.»
Μετάφραση: (από Νίκο Καζαντζάκη και Ιωάννη Καρκιδή)
Μαζί δυο άνεμοι πώς ξεσήκωσαν φυσώντας απ᾿ τη Θράκη, πονέντες και βοριάς, το πέλαγο το ψαροθρόφο, ξάφνου χιμώντας πάνω του, και γρήγορα μαυρίζοντας το κύμα πυργώθη, και σωρός στοιβάχτηκαν στο ακρόγιαλο τα φύκια' παρόμοια κι η καρδιά σπαράζουνταν στων Αχαιών τα στήθη.
Επίσης ο Αριστοτέλης αναφέρει στο βιβλίο του Περί τα Ζώα Ιστοριών «οἷον ἐν χιόνι τῇ παλαιᾷ. Γίνεται δ' ἡ παλαιὰ ἐρυθροτέρα» που είναι αναφορά στο Watermelon snow που οφείλεται σε πληθυσμιακή έκρηξη του φύκους Chlamydomonas nivalis που ζει στο χιόνι και διαθέτει μια κόκκινη χρωστική ουσία την ασταξανθίνη. Στις μέρες μας, η αναζήτηση για ελληνική μικροχλωρίδα συμπεριλαμβανομένων των κυανοβακτηρίων ξεκίνησε τον 20ο αι. με τις μελέτες φυκών γλυκών νερών από τους Stanković (1931) και Skuja (1937). Τις επόμενες δύο δεκαετίες δεν εντοπίζεται κάποια δημοσιευμένη εργασία για μικροχλωρίδα μέχρι τις έρευνες λιμνών του Αναγνωστίδη (1956). Κατά την διάρκεια των 60΄ ο Κωνσταντίνος Αναγνωστίδης έκανε δύο εκτεταμένες μελέτες στα κυανοβακτήρια θερμοπηγών και σε περιοχές με μεγάλες ποσότητες θείου σε περιβάλλοντα θαλάσσια αλλά και γλυκών νερών στην Ελλάδα. Αυτές οι ταξινομικές μελέτες αποκάλυψαν την ύπαρξη πάρα πολλών κυανοβακτηρίων, κάτι το οποίο οδήγησε στην περιγραφή αρκετών νέων ειδών, τα οποία αργότερα εντάχθηκαν στο μοντέρνο σύστημα ταξινόμησης των κυανοβακτηρίων. Η συστηματική αναζήτηση στα κυανοβακτήρια στον ελλαδικό χώρο ξεκίνησε κατά τα 80’ όπου υπάρχουν αρκετές δημοσιεύσεις για την ταξινόμηση των κυανοβακτηρίων, τη μορφολογία των ειδών και τη δυναμική των πληθυσμών τους. Πιο πρόσφατες μελέτες στην ποικιλία των κυανοβακτηρίων στην Ελλάδα περιλαμβάνουν πολυφασικές και πειθαρχειμένες μελέτες για τα γένη των κυανοβακτηρίων, τους φυσικούς πληθυσμούς, καθώς και της οικολογίας τους. Παρ’ όλες τις πληροφορίες που είχαμε μέχρι και το 2016 δεν είχαμε πουθενά συγκεντρωμένα όλα τα είδη που είχαν καταγραφεί στον Ελλαδικό χώρο επισήμων, όταν μια ερευνητική ομάδα του τμήματος Βιολογίας του Αριστοτελείου πανεπιστημίου Θεσσαλονίκης αποτελούμενη από τους: Σπύρος Γκέλης, Ιορδάνης Ουραλλίδης, Μάνθος Πάνου, Νίκος Παππάς, κατέγραψε και δημοσίευσε σε μια λίστα όλη την μέχρι τότε πληροφορία.Συγκεντρωτικά τα είδη κυανοβακτηρίων στην Ελλάδα που είναι γνωστά και δημοσιευμένα σε λίστα μέχρι και σήμερα είναι 543. Τα είδη αυτά που ταξινομούνται σε 130 γένη και 41 οικογένειες και 8 τάξεις. Σε αυτή τη λίστα δεν περιλαμβάνονται διάφορα άλλα είδη που η ταξινόμηση τους είναι ιδιαίτερη, 85 στον αριθμό τα οποία βρέθηκαν σε κόλπους καταρράκτες σπηλιές και λίμνες. Αξίζει να σημειωθεί ότι 18 είδη της λίστας έχουν περιγραφεί μόνο σε ελληνικά περιβάλλοντα καθιστώντας τα ενδημικά και μάλιστα σε σπανίως μελετημένα περιβάλλοντα καταδεικνύοντας την ανάγκη για περαιτέρω μελέτη αυτών. Επιπλέον αναφέρονται διάφορα νέα είδη τα οποία απομονώθηκαν από θαλάσσιους σπόγγους στη Αιγαίο πέλαγος επιβεβαιώνοντας την ανάγκη για περαιτέρω και πιο διεξοδική μελέτη του θέματος, ακόμα και στα πιο απίθανα περιβάλλοντα.
Ο όρος «metabolome» εισήχθη από τον Oliver [14] και αντιπροσωπεύει το σύνολο των μικρών μορίων, ήτοι των μεταβολιτών ενός οργανισμού, οργάνου, ιστού ή κυττάρου. Αυτά τα μικρά μόρια μπορούν να εντοπισθούν μέσω της «Μεταβολομικής» ανάλυσης. Η Μεταβολομική είναι η συστηματική ανάλυση του συνόλου των προϊόντων του μεταβολισμού ενός οργανισμού, δηλαδή, επικεντρώνεται στο εκτενές δίκτυο βιοχημικών αντιδράσεων των κυττάρων ή ιστών, γεγονός που την καθιστά πολύ δυναμική προσέγγιση [15] .
Η ραγδαία εξέλιξη αναλυτικών τεχνολογιών, όπως αέρια (GC) και υγρή (LC) χρωματογραφία, υγρή χρωματογραφία υψηλής πίεσης HPLC και τριχοειδική ηλεκτροφόρηση CE σε συνδυασμό με φασματοσκοπία μάζας MS και φασματοσκοπία μαγνητικού συντονισμού NMR επέτρεψε την εκτεταμένη ανάλυση των μεταβολιτών και πολύπλοκων μεταβολικών μονοπατιών σε σύνθετα βιολογικά δείγματα [16]. Η Μεταβολομική ανάλυση περιλαμβάνει διαφορετικές προσεγγίσεις ανάλογα με τον αρχικό σκοπό μιας πειραματικής μελέτης, βασικότερες εκ των οποίων είναι η «στοχευμένη μεταβολομική ανάλυση» όπου εξετάζονται επιλεγμένες ομάδες μεταβολιτών χρησιμοποιώντας ειδικές τεχνικές διαχωρισμού/ανίχνευσης, η «δημιουργία προφίλ μεταβολιτών» που είναι η ταυτοποίηση και ο ποσοτικός προσδιορισμός των μεταβολιτών που υπάρχουν σε έναν οργανισμό, η «δημιουργία αποτυπωμάτων μεταβολιτών» που είναι μη στοχευμένη, υψηλής απόδοσης ποιοτική διαλογή μεταβολικών συστατικών ενός κυττάρου που αποσκοπεί κυρίως στην σύγκριση δειγμάτων και «ανάλυση μεταβολικής ροής (MFA)» όπου εφαρμόζεται επισήμανση με ισότοπα για να αποκαλυφθούν τα μονοπάτια σε ένα κύτταρο ή οργανισμό [17][18][19][20]. Οι παραπάνω προσεγγίσεις έχουν εφαρμοστεί στην μελέτη του μεταβολισμού των κυανοβακτηρίων. Η έναρξη δημοσιεύσεων για τη μελέτη των κυανοβακτηρίων με τις τεχνολογίες “omics” έγινε το 1996 με την αλληλούχιση του γονιδιώματος του ευρύτατα χρησιμοποιούμενου στελέχους Synechocystis sp. PCC 6803 [21]. Ακολούθησαν μεταγραφωμικές αναλύσεις με την τεχνολογία των μικροσυστοιχιών ή πιο πρόσφατα με αλληλούχιση RNA (RNAseq) οι οποίες συνδυάζονται πλέον με πρωτεομικές μελέτες [19]. Μολονότι η μεταβολομική μελέτη των κυανοβακτηρίων εφαρμόζεται μόλις τα τελευταία χρόνια, η κατανόηση του μεταβολισμού τoυς είναι απαραίτητη καθώς τα κυανοβακτήρια θεωρούνται πλούσια πηγή δευτερογενών μεταβολιτών με πιθανές βιοτεχνολογικές εφαρμογές στον τομέα της φαρμακολογίας και, έτσι, αποτελούν αντικείμενο έρευνας βιοϊατρικών προϊόντων/φαρμάκων και, επιπλέον, είναι σημαντικοί υποψήφιοι για την παραγωγή εμπορεύσιμων βιοκαυσίμων [22][23][19].
Εφαρμογές στοχευμένης μεταβολομικής ανάλυσης στα κυανοβακτήρια στοχεύουν στην ανάλυση μεταβολιτών που συμμετέχουν σε κοινές ή προκαθορισμένες διεργασίες. Παράδειγμα αποτελεί η ανάλυση των τοξινών των κυανοβακτηρίων σε περιβαλλοντικά δείγματα. Για τον σκοπό αυτόν χρησιμοποιούνται η υγρή χρωματογραφία υψηλής πίεσης HPLC και η υγρή χρωματογραφία LC σε συνδυασμό με τη φασματοσκοπία μάζας MS για την ποσοτικοποίηση βιοδραστικών ουσιών (κυκλικά πεπτίδια, αλκαλοειδή κ.α.). Αυτά τα αποτελέσματα θα συμβάλλουν στην δημιουργία αποτελεσματικότερων σχεδίων ελέγχου της ποιότητας του πόσιμου νερού [19].
Ο προσδιορισμός χαρακτηριστικών μεταβολικών μοτίβων σε κύτταρα στελεχών κυανοβακτηρίων αποσκοπεί στην πλήρη κατανόηση του μεταβολισμού των κυανοβακτηρίων και όχι μόνο. Για παράδειγμα, η σύσταση των λιπαρών οξέων χρησιμοποιείται ως δείκτης για την ταξινόμηση μικροβίων τα λιπαρά οξέα μετατρέπονται σε μεθυλ-εστέρες που αναλύονται με GC/MS. Τα αποτελέσματα της ανάλυσης της ολικής σύστασης του κυττάρου σε λιπαρά οξέα αξιοποιούνται στην ταξινόμηση ειδών διάφορων γενών κυανοβακτηρίων [19]. Επιπλέον, ο προσδιορισμός της απόλυτης συγκέντρωσης μεταβολιτών για την δημιουργία μεταβολικού προφίλ συγκεκριμένων στελεχών κυανοβακτηρίων επιτρέπει όχι μόνο την σύγκριση μεταξύ στελεχών αλλά και τον έλεγχο αλλαγών στον κυτταρικό μεταβολισμό στις διαδικασίες μεταβολικής μηχανικής [22].
Η τεχνολογία των μεταβολικών αποτυπωμάτων στα κυανοβακτήρια χρησιμοποιείται κυρίως στον χαρακτηρισμό αλλαγών κάτω από διαφορετικές περιβαλλοντικές συνθήκες ή σε διαφορετικό γενετικό υπόβαθρο. Για παράδειγμα, έχουν χαρακτηριστεί μεταβολικά αποτυπώματα μετά από προσθήκη γλυκόζης στον κύριο μεταβολισμό του στελέχους Synechocystis PCC 6803, χρησιμοποιώντας υγρή χρωματογραφία σε συνδυασμό με φασματοσκοπία μάζας (LC/MS) και βρέθηκε ότι η αύξηση της γλυκόζης σε συνθήκες φωτός είναι τοξική για το κυανοβακτήριο [24][19]. Επιπλέον, οι προσπάθειες δημιουργίας μεταβολικών αποτυπωμάτων με τον διαχωρισμό μεταβολιτών μικρού μοριακού βάρους αποκαλύπτουν και πολλά χαρακτηριστικά από άγνωστους μέχρι στιγμής μεταβολίτες. Για την εξακρίβωση των μεταβολικών τους μοτίβων κρίνεται απαραίτητος ο συνδυασμός διαφορετικών μεταβολικών τεχνολογιών και εκτενείς στατιστικές αναλύσεις. Αυτές οι άγνωστες μέχρι στιγμής ενώσεις μπορεί να σχετίζονται με φυσιολογικά φαινόμενα που είναι υπό μελέτη, επομένως μπορούν να αρχειοθετηθούν και να ενσωματωθούν σε βάσεις δεδομένων ως ενώσεις που δεν έχουν προσδιοριστεί ακόμη και να χρησιμοποιούνται ως δείκτες [19].
Τα κυανοβακτήρια (Cyanobacteria) αποτελούν φωτοσυνθετικούς οργανισμούς που παλαιότερα κατά την κλασική ταξινόμηση φέρονταν με το όνομα «κυανοφύκη». Σήμερα θεωρούνται ιδιαίτερη συνομοταξία στην επικράτεια των Βακτηρίων. Τα κυανοβακτήρια αποτελούν ομάδα φωτοσυνθετικών προκαρυωτικών οργανισμών, που θεωρούνται ικανά σε οξυγονογενή ή οξυγονούχο φωτοσύνθεση.
Η αρχική τους ονομασία «κυανoφύκη» (blue-green algae στη διεθνή ορολογία) οφειλόταν κυρίως λόγω του χρώματος που παρουσίαζαν πολλά είδη εξ αυτών και που προερχόταν από την παρουσία της βιλιπρωτεΐνης που χαρακτηρίζει τις χρωστικές τόσο των οργανισμών αυτών όσο και των ερυθρών φυκών ή ροδοφύτων, και ειδικότερα της φυκοκυανίνης. Όλα τα μέλη της συνομοταξίας αυτής περιέχουν χλωροφύλλη α, εκτός από τα γένη Πρόχλωρον (Prochloron) και Προχλωροκόκκος (Prochlorococcus) που περιέχουν επίσης και χλωροφύλλη β.
Τα κυανοβακτήρια θεωρούνται ως πιθανοί πρώτοι οργανισμοί στη Γη οι οποίοι παρείχαν οξυγόνο με φωτοσύνθεση. Υπάρχουν αποδείξεις, από απολιθώματα του είδους αυτών, περί της ύπαρξής τους πριν από 3,5 δισεκατομμύρια χρόνια. Το δε κυτταρικό τοίχωμα αυτών είναι ανάλογο με εκείνο των «αρνητικών κατά Gram» βακτηρίων, οι δε φωτεινές αντιδράσεις της φωτοσύνθεσης γίνονται μέσα στο σύστημα της θυλακοειδούς μεμβράνης του κυττάρου. Άλλες δομές του κυττάρου τους είναι διάφορα πολυφωσφορικά σωμάτια που παρατηρούνται σαν απόθεμα φωσφορικών αλάτων, καθώς και τα κενοτόπια που πληρούνται με αέριο και βοηθούν στη πλευστότητα αυτών. Μερικά δε εξ αυτών είναι μονοκύτταρα ενώ άλλα είναι νηματώδη. Αναπαράγονται είτε με ένωση είτε με θραυσματοποίηση.
Τα κυανοβακτήρια βρίσκονται σχεδόν παντού σε ξηρά και θάλασσα και ειδικότερα σε χώρους με άπλετο φωτισμό. Μερικά είδη αυτών ζουν σε τελείως αφιλόξενα περιβάλλοντα μέρη, όπως ακόμη και σε θερμοπηγές όπου η θερμοκρασία υπερβαίνει τους +85 °C. Τα κυανοβακτήρια ευθύνονται σ΄ ένα μεγάλο μέρος της εξέλιξης του φωτοσυνθετικού οξυγόνου ιδιαίτερα στους ωκεανούς και συμβάλουν κατά πολύ στη δέσμευση του διοξειδίου του άνθρακα, κατά τις ανεξάρτητες του φωτός (σκοτεινές) αντιδράσεις καθώς και του αζώτου.
Πολλές φορές συγκεντρώνονται σε μεγάλους αριθμούς κατόπιν υπερβολικής ανάπτυξης σε επιφάνειες λιμνών και υδατοδεξαμενών όπου διακρίνονται τακτικότερα σαν πράσινοι επιπλέοντες κόκκοι. Αυτές όμως οι μεγάλες συγκεντρώσεις μπορεί και να απελευθερώσουν τοξίνες, γνωστές ως κυανοτοξίνες.
